
S. G. Moharram
National Institute of Oceanography and Fisheries, Alexandria, Egypt
F. A. Raky
National Institute of Oceanography and Fisheries of Kanater el Khairia, Egypt
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 21 | Page No.: 3742-3751







ABSTRACT
Four week-old Red tilapia fry with an average weight of 1.9 g were reared in saline water (36%) for 190 days on 6 formulated iscaloric diet of protein level (28%). The experiment was conducted in glass aquaria. Six diets were formulated including a control diet based on fish meal (diet 1) and fish silage (diet 2) and soybean (diet 3). The other 3 based diets supplemented with 35% fish meal and 60% fish silage (diet 4), 35% fish meal +65% soybean diet (5) and 50% fish silage +50% soybean (diet 6). There were six treatments in duplicates. The results indicate that supplementation with 65% fish silage let to a significant (p<0.05) growth performance (final weight gain 152.2±1.1 g, SGR 1.08 and daily gain 0.80±0.06 g day-1) and efficient feed utilization (FCR 1.9, PER 2) comparable to other diets. Complete replacement with soybean resulted a significant (p>0.05) low growth performance (weight gain 2.43±0.06 g, SGR 0.65 and 0.41±0.01 g daily weight gain) and less efficient in feed utilization (FCR 2.9 and PER 1.2). The present study also investigated the reproductive performance of red tilapia in sea water. The six diets were fed to red tilapia fries. The first spawning of the brood stock of fish groups occurred at almost the same age. There was a significant (p<0.05) difference in the absolute fecundity fertilization and hatching rate of fertilized eggs among fish fed soybean and the other diets. While no significant difference in the spawning intervals, gonad maturation, egg size, were resulted.
PDF Abstract XML References Citation
How to cite this article
S. G. Moharram and F. A. Raky, 2007. Effect of Varying Dietary Protein Sources on Growth and Spawning Performance and Gonad Maturation of Red Tilapia Reard in Sea Water. Pakistan Journal of Biological Sciences, 10: 3742-3751.
DOI: 10.3923/pjbs.2007.3742.3751
URL: https://scialert.net/abstract/?doi=pjbs.2007.3742.3751
DOI: 10.3923/pjbs.2007.3742.3751
URL: https://scialert.net/abstract/?doi=pjbs.2007.3742.3751
INTRODUCTION
Tilapia is the second most important formed fish group in the world, only surpassed by carps. Tilapia culture in being practiced in most of the tropical, subtropical and temperate regions and has received great attention in recent years.
Many tilapia species are euryhaline and can grow in saline water after acclimation (Suresh and Lin, 1992). Varieties of red tilapia have been successfully cultured in saline water (Watanabe, 1991).
Several studies have been conducted to evaluate the in corporation of different unconventional animal and plant proteins and energy sources for farmed tilapia with varying results (El-Sayed, 1990).
Tilapia has shown a high degree of plasticity in growth and reproduction trails. A comparative analysis across African lakes suggested a negative correlation between the condition factor of fish and the onset of maturation (Lorenzen, 2000). Food supply (quality and quantity) may be expected to have a strong influence on reproduction of tilapia (Brummett, 1995).
Red hybrid tilapia are gaining popularity among culturists due to their resemblance to premium marine species such as sea bream Chryso phrys major and red snapper Lutjanus campechanus and excellent growth and feed conversion rates in freshwater (Liao and Chen, 1983).
The feasibility of rearing red hybrid tilapia in brackish and seawater was first studied by Liao and Chang (1983) who reported good growth of Taiwanese red tilapia O. mossambicusxO. niloticus at salinities of 17 and 37%. Seawater-rearing studies of Taiwanese red tilapia in Kuwait (Hopkins et al., 1985) showed that survival at 38-41% was impaired at water temperatures below 24°C.
Feed represents the single largest expenditure in intensive culture operations and protein is the most expensive component in diets for aquatic species. Fish meal constitutes the major portion of the protein used in most commercial feeds for high value carnivorous fish and omnivorous fish species and marine shrimp. According to Chamberlain and Barlow (2000) fish meal usage within aquafeeds is expected to increase by 50%.
The steady increase of aquaculture production over the last two decades has resulted in an increased utilization of fish meal dominant ingredient used in the production of aquaculture feeds (New and Wijkstrom, 2002). Assuming that fish and crustacean aquaculture production will increase, this means that about 20% of the total world supply of fish meal would be consumed within aquafeeds (New, 1999). Moreover, fish meal use will likely increase in pet foods and specialty livestock feeds and direct utilization in the human diets due to the tremendous increase in the world population (Rumsey, 1993). Therefore, future fish meal prices and hence the costs of fish production will probably be higher than at present unless suitable inexpensive alternative sources of protein, which are of constantly high quality, are found and/or developed (Tacon, 1996).
The present study aimed to investigate the effect of different dietary protein sources on growth performance, sexual maturation and the reproduction performance of red tilapia under laboratory conditions. The spawning performance of brood stock (including first spawning, spawning periodicity, sexual maturation and fecundity), egg size, hatchability rate of fertilized egg were investigated.
MATERIALS AND METHODS
Fish and culture facilities: This experiment was carried out at the National Institute of Oceanography and Fisheries Alexandria (Egypt). Red tilapia fries were obtained from the marine hatchery Kilo 21 at Alexandria Government, mean body weight was 1.93±0.26 g. The fish were stocked into 300 L fiberglass tanks in a closed, recalculating indoor system as an acclimatizing period is about 15 days from November to the start of the experiment at December 2005. Red tilapia were divided into 6 dietary groups. The fish fry kept at density of 5 fish L-1 during the first 3 weeks in 30 L aquaria, then five male and five female juveniles were randomly selected and reared for the following 190 days in 120 L aquaria. The culture system was provided with a biological filter, aeration through an air compressor and heaters to maintain water temperature at 28°C. Approximately 60% of the water volume was replaced by new saline water daily. Water quality parameters including Dissolved Oxygen (DO) (oxygen meter YSI, model 58), ammonia (NH4-N), nitrates (NO3-N) and nitrites (NO2-N) (Orion Aqua fast, Germany) and pH (pH meter, Jenway, UK), were monitored biweekly.
The average values of these parameters throughout the study period were:
Dissolved oxygen = 7.5-9.2 mg dm-3 pH = 7.5-8.0
Ammonia = < 0.005 mg N-NH4 dm-3 Nitrates = 0.01-0.04 mg N-NO3 dm-3; Nitrites = <0.01 mg N-NO2 dm-3
Fish silage fermentation: Trash tilapia fish was collected from Kanater el Khairia fish farm, Egypt. Fish silage (DFFS) was prepared as the method of Fagbenro et al. (1994).
Culture media of lactobacillus plantarum: Lactobacillus plantarum was obtained from microbiological research center, Faculty of Agriculture, Ain Shams University. it was cultured in a media of MRS broth to activate the bacteria by De-Man et al. (1960).
The culture medium was incubated for 24 h at 37°C, then strain was kept in a refrigerator at 5°C (Harrigan, 1966; Attalla, 2001).
Preparation of soybean meal: Endogenous protease was inhibited according to Wee and Shu (1989) raw soybeans were boiled for one hour and then dried into an oven at 48°C for 20 min then milled and dried.
Experimental diets and feeding regime: Six isonitrogenous (28% crude protein) and isocaloric diets (Table 1) were formulated. Ingredient of those diets were fishmeal, fish silage meal and treated soybean meal, wheat bran, rice bran, oil, vitamins and minerals. Feed was introduced to fish once daily at fixed time at 9 am.
Feed composition analysis: Proximate analysis of the studied diets (Table 2) moisture, crude protein, lipid and ash content was performed as described by Lovell (1977).
Rearing condition of red tilapia: During the experiment water temperature was maintained at 28±05°C by using electric heaters with ambient photoperiod. Fish were fed 6 diets. In the control, the fish were fed on fish meal. The sex ratios were checked when the separation was conducted. Fish were fed for 6 days.
Investigation of spawning: Commencing on the day (100-120) when the fish could be sexed. Eggs were removed from the brooder's buccal cavity and the number of eggs in each spawning was counted. Spawning dates were recorded for the calculation of spawning periodicity (mean number of days taken for an individual fish complete one spawning cycle).
Total fecundity, defined by Rana (1988) as the number of eggs in fresh spawned clutch was determined directly and egg diameter were measured with a microscope containing eye piece micrometer. Moreover, hatchability rate was determined.
For the histological analysis, gonad samples in the breeding season were fixed in Bouin’s solution. Subsequently the specimens were dehydrated in alcohol with increasing concentration, fixed in xylene and left intact for paraffin embedding.
| Table 1: | Composition of experimental diets containing different fish meal replacers for Red tilapia |
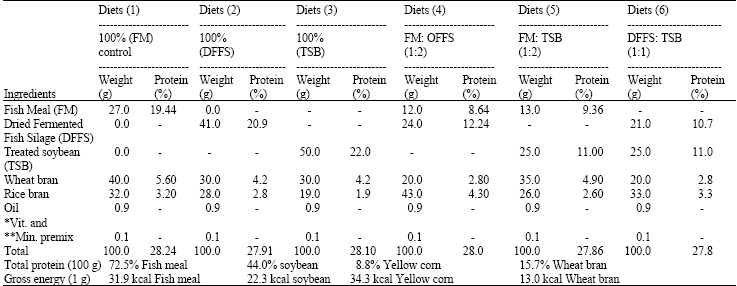 | |
*: Vitamins (premix): Each gram contains Vit A: 4.000.000 IU Vit D3: 7.000.00 IU, Vit H: 3000 mg, Vit K: 6000 mg, Vit B1: 300 mg, Vit B2: 1500 mg, B6, 500 mg, B12: 10 mg, Vit Pantothenic 3000 mg, Folic: 300 mg, Niacin amide: 10000 mg, Biotin: 15 mg; **: Mineral (premix) in mg g-1: Manganese chloride: 30, Iron 10000, Zinc oxide 20000 cupper 1300 senilium: 15000 Cobalt chloride: 30 mg | |
| Table 2: | Chemical composition and energy content of supplementary diets fed for Red tilapia |
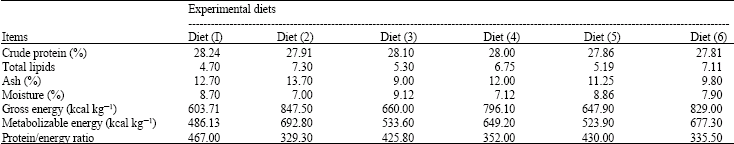 | |
The sections were cut into pieces 8 and 5 μm thick for ovary and testis respectively and stained with the HE (haematoxylin and eosin). Then, serial cross-sections were analyzed for the shape, size, blood flow and the type of germ cells present in gonads. The cell diameters were measured (±1.0 μm) under a microscope using stage micrometer.
Statistical analysis: Data were analyzed statistically using Analysis of Variance (one-way ANOVA) according to Snedecor and Cochran (1980), followed by comparison of treatment means by Duncan's multiple range tests (Duncan, 1955).
RESULTS
Growth performance: Growth performance of red tilapia raised in seawater (36‰) fed on 6 diets which are isocaloric diets containing 28% protein was shown in (Table 3). It is obvious that red tilapia can be grown on 28% protein diets in glass aquaria from small fry (1.3-2.3 g) to adult sizes (152.2±1.1 g).
All growth performance parameters were best in the fish fed on diet 4 and were significantly better than those fed on other diets (p<0.05).
Growth performance in diet 3 was lowest and significantly lower than that in other diets (p>0.05). As the soybean meal content increased, feeding efficiency and protein utilization of the diet decreased (Table 3). In addition, the Feed Conversion Ratio (FCR) in diet 3 was highest and significantly (p<0.05) higher than that in other diets. Protein Efficiency Ratio (PER) (Table 4) was significantly higher (p<0.05) in diets 1, 2 and 4 (2) than at diets 3, 5 and 6 (1.2, 1.6 and 1.5), respectively.
From Table 3 it is obvious that diets 5 and 6 which were supplemented with fish meal and silage improved the growth performance of fish group fed on them.
Statistical analysis displayed significant difference (p<0.05) on the survival of fish after 190 days fed on the different diets (Table 3 and 4).
Table 4 shows the calculated levels of essential amino acids provided by protein of fish meal, silage and soybean meals. It is clear that the percentage of mithionine, lycine and valine were lower in soybean meal. Result indicates that soybean diet deficient in those amino acids lead to reduced growth performance of red tilapia fed on this meal (Table 3).
Carcass composition: Results in Table 2 showed the chemical analysis of fish fed on six diets.
| Table 3: | Growth performance of hybrid red tilapia fed different studied diets |
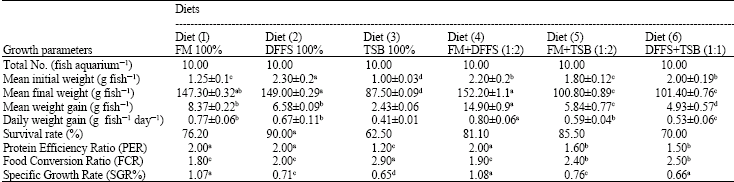 | |
| a, b, c, d = Different alphabets in the same row means significant at 5% | |
| Table 4: | Chemical analysis, energy content and essential amino acids of the experimental ingredient fed to Red tilapia |
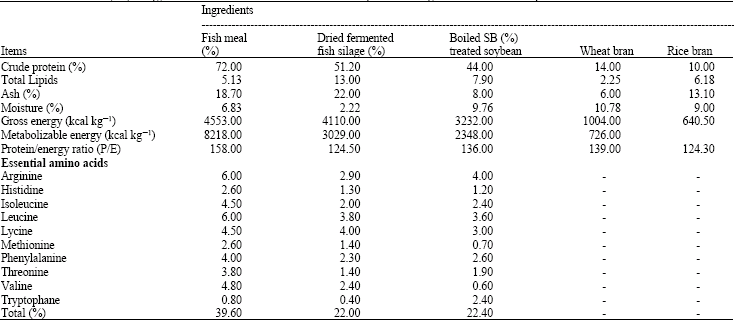 | |
| Table 5: | Spawning performance of red tilapia fed six diets |
 | |
| a, b, c, d = Different alphabets in the same row means significant at 5% | |
The fat content of fish fed on diets 2 and 6 was higher (p<0.05) as compared to those fed other diets. While, ash content in fish caracass decreased (p<0.05) when fish fed on diets 3 and 6 as compared to other diets. The moisture content was higher in diet 3 as compared to other diets.
Spawning performance: Spawning performance of red tilapia females maintained on 6 diets are shown in Table 5.
No significant difference (p>0.05) was found in the onset of first spawning between the six groups, 190±10 day for group 1 and 182±12 days for group 2, while 178±18 for group 4 and 180±12 for groups 5 and 6 and for group 3 it was 174±18 days.
Reproductive status
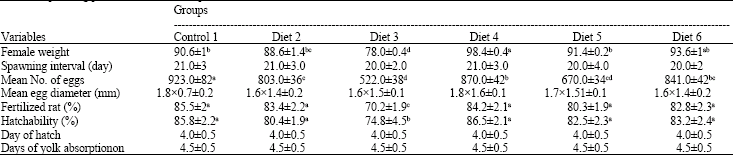
Gonadal maturation: Males were longer and heavier than females of the same age in all groups. Males matured earlier than females: the first mature males were recorded when they were 14 weeks old, whereas the females matured after 18 weeks. In all treatments spawning was initiated when the fish were 20 weeks old. Smallest size at spawning of males and females was 9 cm (13.6 g) and 7.8 cm (8 g), respectively. Both male and female gonads showed presence of 4 maturation stages through the study period.
Testes: The testes were non-zonal, lobules were loosely organized and the zonation was not apparent. The different parts of the testis i.e., anterior, middle and posterior parts showed morphohistological variations in different maturity stages. However, all the three parts of the testis showed similar stages of development in a particular maturity stage. Macroscopic observations of the gonad developmental stages showed that immature males had translucent, non-vascularised, thread like testes. In early maturing testes become enlarge, flat, increase in weight and volume and creamy white in color. While in the developing males, testes enlarge, increase in weight and volume, light pinkish and thicker in size and look more vascular. Blood capillaries become conspicuous. The testes of ripe males appeared swollen milky white and well vascularised and ready released milt under gentle pressure of the abdomen. In spent males the testes became flabby, thin and dull white in color. In Sections of the testes of both immature and resting phase (Fig. 1a), numerous spermatogonia were observed inside the small seminiferous lobules. The spermatogonia were large, spherical cells containing a large round, central nucleus with distinct nucleolus. In mature stages (2 and 3) intense spermatogenesis was seen during this phase. Spermatogonia decreased in number and numerous primary and secondary spermatocytes were visible (Fig. 1b and c). In ripe stage, blood capillaries became conspicuous, the seminiferous lobules were larger in size and full of sperm. Spermatogonia were few and all stages of spermatogenesis can be seen in various lobules (Fig. 1d). In sections taken from spent males large empty areas are apparent and only small numbers of spermatogonia spermatocytes and spermatozoa were present.
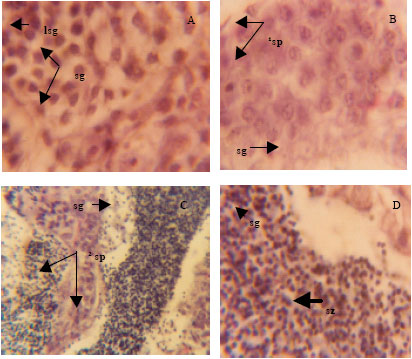
Ovaries: The wall of the ovary was fairly thick during the non-breeding season but became thin and highly vascular during the spawning period. The ovarian lumen was loosely organized and the zonation was not apparent. Immature females had ovaries very small, thin, thread like pale in color, occupying a small part of the body cavity. In early maturing stage (2) ovaries become slightly larger and increase in weight and volume with minute opaque whitish eggs occupied about half of the body cavity. In developing stage (3) ovaries distended occupied, about 2/3 of abdominal cavity with large pale yellow eggs.
 | |
| Fig. 1: | TS of testes of red tilapia showing. (a) Immaturing stage, (b) Maturing stage, (c) Stage 3 and (d) Ripe stage, L: seminefrous lobule, lsg: large spermatogonia with large nucleus (head arrow), sg: small spermatogonia, 1sp: primary spermatocytes, 2sp: secondary spermatocytes, sz: spermatoxoa |
Ripe females had ovary which becomes more enlarged occupying almost entire body cavity, with large number of big, turgid, spherical, translucent, deep yellow ripe ova. Ovary walls became thin almost transparent. Ripped eggs are visible through the ovarian wall. In spent females, ovaries are flaccid, shrinked and sac like, reduced in volume. Microscopic examination of sections of ovary shows that in maturing phase, the ovary contains numerous oocytes in different stages of development (Fig. 2a). Oocytes in Stage 1 to 3 were large in number. Descriptions of each stage of oocyte are as follows; Oocyte 1, this was spherical in shape with strongly basophilic cytoplasm, central nucleus, having 2 or 3 nucleoli; Oocyte 2, there was further increase in the size of the oocyte and basophilic of the cytoplasm. Nucleus enlarged in size and the number of nucleoli increased; Oocyte 3, there was further increase in the size of the oocyte and was distinguished by the appearance of a thin layer of follicular cells around the cytoplasm. A large number of small, clear vacuoles called the yolk vesicles, appeared in the periphery of the ooplasm. The vesicles appeared red in eosin dye. In stage 3, vascular supply increased and the blood capillaries became conspicuous. Also immature oocytes were reduced in number, the oocyte grows further, the yolk vesicles increased in number and fill the entire ooplasm and a vitelline membrane or zona radiata was also clearly visible, between teooplasm and the follicular layer or the zona granulose (Fig. 2b). In ripe ovaries, a large number of oocyte which is characterized by heavy deposition of yolk globules. Migratory nucleus was seen towards the periphery. Some yolk vesicles were pushed towards the periphery of the egg and form cortical alveoli. The yolk granules fused to form larger globules. A thin layer of fibroblasts (theca) was also distinguishable outside the follicular layer (Fig. 2c). In spent phase, the ovary showed atretic and discharged follicles, along with stage 1 and 2 oocyte (Fig. 2d).
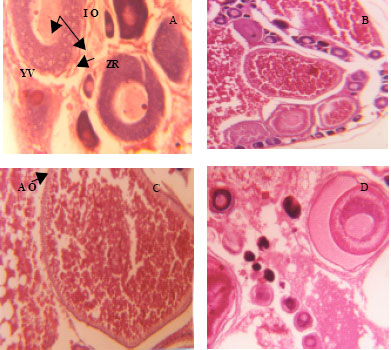
Oocytes development: Microscopic examinations of ovaries showed that oocytes diameter varied from 100μm to 1800 μm. Oocytes frequency distributing show the simultaneous occurrence of oocytes different size indicating that each individual spawns more than once within the breeding cycle (Fig. 3). In addition to oocytes in the initial stages of maturation germ cells with well defined cytoplasm and one or two spherical nucleoli predominated. Mature oocytes were present which were bigger and had rich lipid stores. Lacunae were apparent in the ovaries of spent ones.
 | |
| Fig. 2: | TS of ovary of female red tilapia showing (A) Maturing stage, the ovary contains oocytes in different stages of development. 1, 2I and 3). X 225, (B) stage 3, (C) Ripe stage. X400 (D) spent stage. X 225, AO: Ateritic Oocyte, cy: Cytoplasm., EF: Epithelial Follicle., IO: Immature Oocytes., n: Nucleus, RO: Ripe Oocyte, YG: Yolk Granulose, yv: Yolk Vesicles, ZR: Zona Radiata |
 | |
| Fig. 3: | Showing the ova diameter during spawning season; (a) The banging of spawning season; (b) The middle of spawning season and (c) The end of spawning season |
| Table 6: | Absolute fecundity of red tilapia |
 | |
Fecundity: Absolute fecundity in red tilapia is very variable as shown in Table 5. Absolute fecundity (number of eggs) was significantly higher (p<0.05) in the control group w hile it was lowest at the soybean protein diet (522, Table 5). In this study, the average absolute fecundity varied from 522-923 eggs in the 6 diets and significantly differ (p<0.05). Table 6 showed the absolute fecundity in red tilapia regardless to diets. It was found that the absolute fecundity of the species varied from 470 to 988 eggs per female for a size range of 7.8 to 15 cm (8 to 53.82 g).
Egg and larval quality: From Table 5 it is cleared that no significant differences between the control and other diets in the egg size. A significant difference (p>0.05) was found in fish fed on diet 3 where the fertilization and hatchability rates were 70.2 and 79.8%, respectively.
Sex ratio: The sex ratios checked at the end of the experiment were 2.6:1 female to male respectively for populations of diet 1 (control) and diet 2 while of population of diet 3 it was 1:1.3 (female: male). In populations of diet 4, 5 and 6 the sex ratio was 2:3 female to male. In general the sex ratio of all fish was approximately 1:1.8 female to male (35.7: 64.3%).
DISCUSSION
The growth of tilapia has been found to be highly plastic, responding immediately to changes in resource availability and other factors (Lorenzen, 2000).
Hafedh (1999) showed that tilapia fry, weighing 0.5 g, had the best growth and matured earlier when fed protein levels around 40%. As tilapia increased in age (100-200 g), optimal dietary protein level was shown to decrease to 30%. Twibell and Brown (1998) argue that fish weighing approximately 21 g require a minimum crude protein allowance of 28% for maximum growth and feed efficiency. According to the results of the present study, red tilapia fry (1.3-2.3 g) fed 28% crude protein can be grown and spawn. This result, coincide with those of Watanabe and Wicklund (1993) for this hybrid fry, who showed that red tilapia can be grown on 20-30% crud protein.
The FCR, SGR and PER results showed that fish fed on diet 4 performed significantly better than other group (Table 4). According to the results in Table 4 best growth performance was indicated when fish fed on diet 4. These results, coincide with those of Rose et al. (2002) who found that pacu, Piaractus mesopotamicus fed on diets containing fish silage showed better results than the control diet, which contained fish meal as the only animal protein source.
In the present study, better growth performance was obtained when fish fed on fish silage (diet 2) resembled that of diet 1 and 4 diets. In eel fingerlings, an increase in the specific growth rate, the food conversion efficiency and the protein efficiency ratio has been found when the fish fed isoproteic and isocaloric diets with different percentages (10, 15 and 20%) of fish silage (Gonçalves et al., 1989). Also, juvenile catfish Clarias gariepinus fed on the fish-silage diet showed some significant (p<0.05) differences in mean weight gain, specific growth rate and protein productive value, but feed conversion and protein efficiency ratios were similar (p>0.05) (Fagbenro and Jauncey, 1995). While, Darmawiyanti et al. (2005) reported that replacement fish meal by fish silage in humpback grouper Cromileptes altivelis fingerling diet showed that growth rate and feed conversion of humpback grouper fed the various test diets was not significant different.
Much research has been done to evaluate the nutritional value of soybean meal as a substitute for fish meal. Most studies showed that total replacement of fish meal by soybean meal resulted in reduced growth and feed efficiency. In the present study there were significantly differences in growth performance parameters in fish fed on diet 3. Simultaneously, growth performance decreased with increasing the percentage of soybean in the diets. As the soybean meal content increased, feeding efficiency and protein utilization of the diet decreased (Table 4). The 100% substitution of fish meal with soybean did however result in reduced weight gain (87.5±0.09) and can there-fore be deemed unsuitable for complete Fish meal substitution for this fish species. Red tilapia fed on 100% soybean (diet 3) appeared to grow significantly slower than those fed on fish and fish silage meals. These results are consistent with data of Fagbenro and Davis (2004) who reported that at 100% replacement of fish meal with soybean growth and feed utilization were lower in African catfish clarias gariepinus compared with other treatments. Also, Hernández et al. (2007) studied the possible use of soybean meal in sharp snout sea bream Diplodus puntazzo diets by progressively increasing its inclusion level (0, 20, 40 and 60%), they observed that the diet containing the highest level of soybean meal produced the lowest final weight and as the soybean meal content increased, feeding efficiency and protein utilization of the diet decreased.
Essential amino acid deficiency has been widely demonstrated to reduce feed intake, where normal intake values are reached only when the amino acid concentration in the diet meets the requirements of the fish, as reviewed by De la Higuera (2001). Soy meal is the vegetable-protein source most frequently used in practical diets to replace fish meal. Nevertheless, its methionine deficiency for most fish species limits fish meal-protein substitution. Results in Table 3 revealed that the percentage of lucien, methionine and valine in soybean was lower than those of other. With hybrid tilapia (Tilapia nilotica x Tilapia aurea), at a sub-optimum dietary protein level (24%) fish meal can be partially replaced by soybean meal. At the optimum dietary level of protein (32%), replacement of 30% fish meal with soybean meal required the addition of methionine to the level of the control diet to prevent depressed growth and feed efficiency (Shiau et al., 1987).
Complete substitution of fish meal by soybean meal resulted in reduction of weight gain and feed efficiency, but these can be overcome by supplementation with oil, lysine, methionine and vitamins (Shiau et al., 1987).
In the present study, the 100% substitution of fish meal with soybean meal did however result in reduced growth performance (SGR, 0.65, 0.41 daily weight gain and FCR, 2.9, Table 4). These findings are in agreement with Hunge (2004) who reported that red tilapia fed on complete replacement fish meal with soybean without supplementation of methionine resulted low growth performance (SGR 2.3 and 59 daily weight gain and increased FCR, 2.29).
On the contrary, Nyirenda et al. (2000) tested the effect of replacing animal protein (fish meal, meat and bone meal) with soybean meal (10:0, 5:5, 0:10% of diet) as the protein source on growth and feed conversion of Oreochromis karongae. There were no significant differences in Growth Rate (GR), Specific Growth Rate (SGR) and Feed Conversion Ratios (FCR) among the three diets. Bonaldo et al. (2006) determined the effect of three isoproteic and isolipidic extruded diets (crude protein, 47%, crude fat, 20%) containing 0, 18 and 30% of soybean (SBM) for 87 days on growth performance, nutrient utilization Egyptian sole, Solea aegyptiaca. They reported that no differences in performances or nutrient retention were found. On the basis of their results, SBM seems to be a good protein source for Egyptian sole and can be added in the diet up to 30% without any reduction in growth rate.
Spawning was observed in all diets in seawater (36%) in this experiment, which is consistent with the results obtained by Watanabe et al. (1989).
No significant difference was found between the 6 diet groups in term of egg diameter. These results are consistent with data reported by Rana (1988) that within a group of females of the same age class there is no significant relationship between body size and egg size. Similar observation has been made on tilapia nilotica (Al-Hafedh et al., 1999).
Results of the present study demonstrate that smallest size at spawning of males and females was 9 cm (13.6 g) and 7.8 cm (8 g), respectively. This observation is consistent with data recorded by Hafedh et al. (1999). Hatikakoty (2002) reported that female and male O. mossambicus matured at the average length of 7.5 and 12.5 cm, respectively indicating that females mature at smaller size than their male.
In the present study it was observed that absolute fecundity varied from 470 to 988 eggs per female for a size range of 7.8 to 15 cm (8 to 53.82 g). This result is in agreement with Hatikakoty (2002), who reported that the absolute fecundity of O. mossambicus varied from 100 to 850 eggs per female for a size range of 7.6 to 19.9 cm. De Silva and Chandrasoma (1980) found that the fecundity of O. mossambicus varied from 360 to 1,775 eggs per female for fish with weight ranging from 145 to 538 g. In this study, the absolute fecundity of the species varied in the six diets from 522-923 eggs per female (78 to 98.4 g) (Table 5).
According to results in Table 5 it is obvious that there were significant differences in fertilization and hatchability rate in group fed on diet 3. These findings are in agreement with results of Lu and Takeuchi (2004).
The average percentage of red tilapia males in this experiment was 64.3%. El Gamal (1987) reported that sex ratio of red tilapia were skewed consistently toward males, while in a study on reproductive efficiency of red tilapia it was noticed that sex ratio were highly variable (Smith and Phelps 2000).
REFERENCES
- Al-Hafedh, Y.S., A.Q. Siddiqui and M.Y. Al-Saiady, 1999. Effects of dietary protein levels on gonad maturation, size and age at first maturity, fecundity and growth of Nile Tilapia. Aquacult. Int., 7: 319-332.
CrossRefDirect Link - Bonaldo, A., A.J. Roem, A. Pecchini, E. Grilli and P.P. Gatta, 2006. Influence of dietary soybean meal levels on growth, feed utilization and gut histology of Egyptian sole (Solea aegyptiaca) juveniles. Aquaculture, 261: 580-586.
CrossRefDirect Link - Coward, K. and N.R. Bromage, 1999. Spawning frequency, fecundity, egg size and ovarian histology in groups of Tialpia zillii maintained upon two distinct food ration sizes from first-feeding to sexual maturity. Aqua. Living Resour., 12: 11-22.
CrossRefDirect Link - De la Higuera, M., 2001. Effects of Nutritional Factors and Feed Characteristics on Feed Intake. In: Feed Intake in Fish, Houlihan, D., T. Boujard and M. Jobling (Eds.). Blackwell Science, Oxford, pp: 250.
CrossRef - De Silva, S.S. and J. Chandrasoma, 1980. Reproductive biology of Sarotherodon mossambicus, an introduced species, in an ancient man-made lake in Sri Lanka. Environ. Biol. Fishes, 5: 253-259.
Direct Link - El-Sayed, A.F.M., 1992. Effects of substituting fish meal with Azolla pinnata in practical diets for fingerling and adult Nile tilapia, Oreochromis niloticus (L.). Aquacult. Res., 23: 167-173.
CrossRefDirect Link - Fagbenro, A.O., K. Jauncey and G. Haylor, 1994. Nutritive value of diets containing dried lactic acid fermented fish silage and soybean meal for juvenile Oreochromis niloticus and larias gariepinus. Aquat. Liv. Resour., 7: 79-85.
CrossRefDirect Link - Fagbenro, A.O. and K. Jauncey, 1995. Growth and protein utilization by juvenile catfish (Clarias gariepinus) fed dry diets containing co-dried lactic-acid-fermented fish-silage and protein feedstuffs. J. Bioresour. Technol., 51: 29-35.
CrossRef - Gon�alves, J.E., S. Sofia, S. Victor, I.B. Pereira and C. Jo�o, 1989. The use of fish Silage as an ingredient for eel fingerling nutrition. Aquculture, 80: 135-146.
CrossRef - Harrigan, W.F., 1966. The nutritional requirements and biochemical reactions of Corybacterium bovis. J. Applied Bact., 29: 380-393.
CrossRefDirect Link - Nyirenda, M., E. Mwabumba, E. Kaunda and J. Sales, 2000. Effect of substituting animal protein sources with soybean meal in diets of Oreochromis karongae (Trewavas 1941). Naga, 23: 13-15.
Direct Link - Rose, M.V., J.C. Dalton, M. Elizabete and V. Macedo, 2002. Growth rate of Pacu, Piaractus mesopotamicus, fingerlings fed diets containing fish silage as replacment of fish meal. J. Applied Aquacult., 12: 77-88.
Direct Link - Snedecor, G.W. and W.G. Cochran, 1980. Statistical Methods. 7th Edn., Iowa State University Press, Iowa, USA., ISBN-10: 0813815606, Pages: 507.
Direct Link - Hernandez, M.D., F.J. Martinez, M. Jover and B.G. Garcia, 2007. Effects of partial replacement of fishmeal by soybean meal in sharpsnout seabream (Diplodus puntazzo) diet. Aquaculture, 263: 159-167.
CrossRef
S.K.Prabhakar Reply
This paper is unique and usefull for research work.