
Zamani Zahra
Department of Biochemistry, Pasteur Institute of Iran, Tehran, Iran
Razavi Mohammad Reza
Department of Biochemistry, Pasteur Institute of Iran, Tehran, Iran
Assmar Mehdi
Department of Parasitology, Pasteur Institute of Iran, Tehran, Iran
Sadeghi Sedigheh
Department of Biochemistry, Pasteur Institute of Iran, Tehran, Iran
Pourfallah Fatemeh
Department of Biochemistry, Pasteur Institute of Iran, Tehran, Iran
Nasoohi Nikoo
Department of Biochemistry, Pasteur Institute of Iran, Tehran, Iran
Sheibani Ashraf
Al-Zahra University, Tehran, Iran
Raisi Mohammad
Center for Control of Infectious Diseases, Ministry of Health, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 3 | Page No.: 368-372







ABSTRACT
Plasmodium falciparum merozoite surface protein-1 (MSP-1) shows extensive antigenic diversity. This is due to the presence of seven variable blocks, five semi-conserved and also five conserved blocks. The variable blocks in the MSP-1 gene are principally dimorphic, displaying either K1 or MAD20 type; except for the block 2 region which is represented by three alleles, an RO33 type in addition to the other two. Allelic diversity is reported to be generated by intra-genic recombination between the variable blocks. A study of allelic variation of MSP-1 gene in Plasmodium falciparum was carried out in the southern province of Sistan Baluchistan in Iran in 2001-2003. Samples were obtained from 30 febrile patients and DNA was extracted and association types between blocks 2 and 6 was identified on each block using specific primers and compared with those from Vietnam, Brazil and Africa. The association types obtained, were similar though less in number than the ones from Vietnam, but more than those from Africa and Brazil.
PDF Abstract XML References Citation
How to cite this article
Zamani Zahra, Razavi Mohammad Reza, Assmar Mehdi, Sadeghi Sedigheh, Pourfallah Fatemeh, Nasoohi Nikoo, Sheibani Ashraf and Raisi Mohammad, 2007. A Preliminary Study of Genetic Diversity of MSP-1 Types in Plasmodium falciparum in Southern Province of Sistan Baluchistan of Iran. Pakistan Journal of Biological Sciences, 10: 368-372.
DOI: 10.3923/pjbs.2007.368.372
URL: https://scialert.net/abstract/?doi=pjbs.2007.368.372
DOI: 10.3923/pjbs.2007.368.372
URL: https://scialert.net/abstract/?doi=pjbs.2007.368.372
INTRODUCTION
Malaria causes over 300 million cases and 2 million deaths annually world wide (WHO, 1993) Plasmodium falciparum is one of the main causes of cerebral malaria and the most pathogenic protozoan parasite (Girard et al., 1988). Increased insecticide resistance by the mosquito and drug resistance by the parasite itself has made vaccine preparation an urgency, (Cowman et al., 2000). One of the vaccine candidates for the blood stage parasites is the 190-200 kD major glycoprotein merozoite surface protein-1 (MSP-1) which after proteolytic processing, leaves only a 19 kD C-terminal fragment remains anchored on the merozoite surface during invasion (Blackman et al., 1992). An immune response to MSP1 is expected to stop merozoite invasion of erythrocytes, the developmental stage of the parasite that causes clinical malaria (Cooper et al., 1992; Pirson and Perkin, 1985; Riley et al., 1992; Riley, 1996). The protein however, shows extensive polymorphism, but is very strongly associated with immunity to the disease (Diggs et al., 1993).
An alternative approach could be a multivalent vaccine based on polymorphic antigens whose composition can be changed regularly to match locally prevalent antigenic variants. This could be an alternative to those made of highly conserved but poorly immunogenic antigens (Anderson et al., 1997).
Tanabe et al. (1987) compared different sequences and described 7 variable regions in the MSP1 gene interspersed with conserved or semi-conserved regions. The 19 kD C-terminus region corresponds approximately to thee conserved block 17 (Fig. 1) which is a major target of naturally acquired anti-malarial immunity (Tanabe et al., 1987). There are two basic versions of each block named after the representative isolates K1 and MAD20 (Certa et al., 1987). The only known exception to the allelic dimorphism occurs in block 2 which has a third version named RO33 (Peterson et al., 1988). Most allelic diversity is generated by recombination near the 5` end of the gene and the variations in the tripeptide repeats found in the MAD20 and K1 version of block 2. This block of MSP1 has been defined as a principal target of antibodies associated with clinical immunity in Amazonian patients (Da Silveira et al., 1999).
Almost all possible combinations of PfMSP1 variable blocks, are found in clinical isolates (Da Silveira et al., 1999). A similar study has been carried out in Brazil, Tanzania and Southern Vietnam and it was seen that some combinations and gene types from different continents (Kaneko et al., 1997) are significantly more prevalent than expected by random assortment of allelic types (Ferreira et al., 2003).
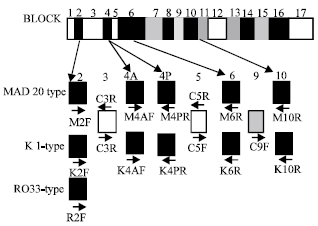 | |
| Fig. 1: | Primers for determination of allelic types in variable blocks of the plasmoium falciparum MSP1 gene. Conserved, semiconserved and variable blocks of the gene are shown as white, gray and black boxes, respectively. Block numbers are after Tanabe et al. (1987). Location and directions of primers are indicated by arrows |
Using the PCR strategy developed by Kaneko et al. (1997), we carried out a study of the patterns of allelic diversity in Iranian patients infected by P. falciparum between the years 2000-2002 from Sistan Blauchistan State.
MATERIALS AND METHODS
Sample preparation: 2 ml of blood was obtained from 30 willing febrile patients confirmed as being infected with Plasmodium falciparum by microscopic examination of Gimesa stained thick blood smears. The patients were residing in nearby cities in the Sistan Baluchistan province in south-east Iran. Information revealing their personal data, details of drug therapy and recent travels to neighboring countries were noted in a questionnaire.
DNA extraction: 1.5 mL blood was centrifuged (1200 rpm for 5 min) at 4°C. The precipitate was washed with RPMI or PBS. Erythrocytes were initially lysed in 700 μL PBS buffer containing saponin on ice for 5 min. Cells were then centrifuged at 10,000 g for 5 min and washed twice with 1 mL PBS. Lysis was carried out in lysis buffer (50 mM tris (pH 7.8), 50 mM EDTA, 10 mM NaCl, 1% triton X-100 ) and 15 μL proteinase K overnight at 60°C. Phenol chloroform extraction was done according to the method of Sambrook et al. (1989). DNA was dissolved in 120 μL deionized DW. The entire experiment was carried out on ice.
PCR amplification: 35 PCR cycles (each one 3 min at 94°C, 45 sec at 91°C, 45 sec at 50°C and 45 sec at 70°C and 7 min at 70°C) in a thermal cycler (Eppendorf). PCR amplification was performed in a 25 μL reaction mixture containing 1 μL each of forward and reverse primers designed by Tanabe et al. (1987), 1 μm of dNTP, 1 unit of DNA polymerase (Roche ), 2.5 μL of PCR buffer and 1 μL of DNA were prepared from the infected erythrocytes Every PCR assay included a tube without template as negative control. PCR products were electophoressed on 0.8 to 2% agarose gel (depending on predicted size of PCR products), in 0.5X Tris-borate EDTA buffer and documented with gel doc (Mitsubishi).
Determination of MSP-1 association type by PCR: Sequences of oligonucleotide primers and PCR protocols was used to type the blocks as described by Tanabe et al. (1989).
RESULTS
The common primer between block 3 and 5 was detected in all samples (Fig. 2a) as a heavy band of 500 bp. Allelic family typing of block 2 with one of the 3 allele specific primers (K2F, M2F, R2F) and a conserved primer (C3R) (Tanabe et al., 1987) yielded specific bands of predicted sizes (Fig. 2a). Specific amplification was also observed for typing of block 6 with a conserved primer C5F and either of the two allele specific primers (K6R and M6R) (Fig. 2c). All the three variable types of block 2, K1, MAD20 and RO33 type were detected (Fig. 2b). Nested PCR of block 4 once again demonstrated the presence of both K1 and MAD20 types (Fig. 2d).
In total, 98 MSP-1 association type clones were determined from 30 blood samples. Twelve out of 30 samples exhibited two different association type clones and more than 3 association type clones were seen in another 12 isolates including those containing as many as 8 clones. Thus, 24/30 isolates (80%) contained multiple association type clones (Fig. 3). The average number of MSP-1 association type clones per patient was 3.26 in Sistan Baluchistan in 2001-2003.
Frequency of MSP-1 association types: The analysis of 98 MSP-1 association type revealed 19 distinct types. The frequency distribution of the expected 24 association types was shown in the histogram (Fig. 4). As frequency is expressed only for MSP-1 association type clones and not parasite clones, it is probable that the same MSP-1 association type clone is shared by different parasite clones.
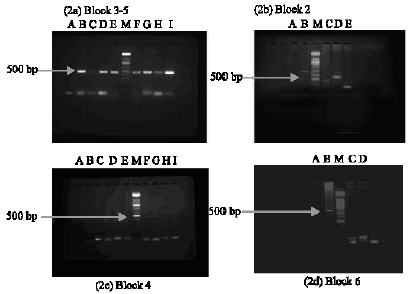 | |
| Fig. 2: | PCR amplified fragment of the Plasmodium falciparum MSP-1 gene variable blocks, 2, 3-5, 6 and nested PCR of block 4 using specific primers analyzed by 2% agarose gel electrophoresis. MW markers are 100 bp ladders containing a mixture of 15 double stranded DNA ranging from 100 to 1500 bp containing an additional 2642 bp band (Roche). (2a) Lane A is without template and lane M contains MW markers. And the rest show common primers for analysis of blocks 3-5; 500 bp fragment amplified by common primers (2b) shows variable block 2-3; Lane A, without template, lane B positive control, lane C RO33 at 270 bp, lane D, K1 at 264- 269 bp and lane E shows MAD 20 at 154 bp and lane M, molecular weight marker (2c) Nested PCR using amplified fragment from block 2-5, lane A without template, lane B positive control, lane M molecular weight markers, lane I MAD20/MAD20 97 bp, lane H MAD20/K1 at 94 bp, lane G K1/K1at 80 bp and lane F K1/MAD20 at 84 bp (2d) lane A positive control, lane B MAD20 at 111 bp, lane C K1 at 119 bp, lane D without template and lane M molecular weight markers |
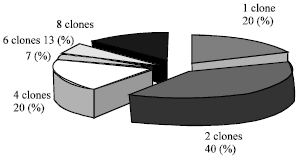 | |
| Fig. 3: | Percentage of MSP-I genotype clones |
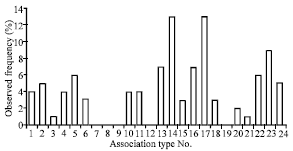 | |
| Fig. 4: | Histogram depicting frequency percentage of MSP1 association types. Frequency distributions of MSP1 association types were compared by χ2 test on 2 x R contingency tables; 5% was employed as the level of significance |
Different kinds of association type numbers combinations of K1, MAD20, RO33 in blocks 2, 4a, 4P and 6.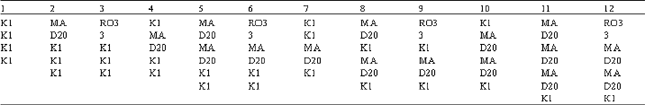 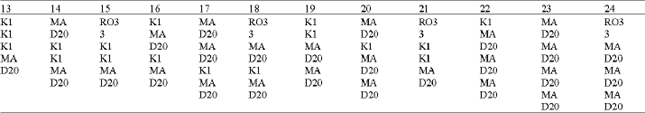 |
Particular association types were predominant in Sistan Baluchistan like type number 17 (with MAD20 type in blocks 2 and 4A, K1 type in block 4P and MAD20 type in block 6) and association type 14 (with MAD20 type in block 2 and block 6 and K1 type in 4A and 4P). Association types having MAD20 type in block 6 existed at a high rate in all isolates (right half of histogram, Fig. 4).
DISCUSSION
During the period of our study from 2001-2003, the total number of malaria cases in Iran dropped from 19,303 to 15,558 cases. In 2001 17,145 P. vivax cases were reported, whereas in 2003, it had dropped to 13,1318 and the P. falciparum cases from 1875 had gone to 2,219 whereas mixed infection dropped to 195 (Communications from the Center for Prevention of Communicable Diseases, Tehran, Iran, 2004 (unpublished)). On the whole, malaria cases had reduced and the downward trend is fortunately continuing.
Allelic variations arise due to different associations in the variable blocks which could be either of K1, MAD20 or RO33 types. The total number of association types reported so far is 24 (Kaneko et al., 1997; Da Silveira et al., 1999; Ferreira et al., 2003) though it is assumed to be much more, as the conserved blocks also show sequential variations. This is demonstrated by 6 different sizes of the K1 allelic types in block 2 which could be due to size polymorphism of the tripeptide repeats, (Jonguwities et al., 1992; Miller et al., 1993; Daubersies et al., 1996; Kaneko et al., 1997), which was also observed in this study.
Earlier studies in Vietnam have reported 21 (n = 186), Brazil have shown 15 (n = 620) and Tanzania 13 (n = 60), whereas we have observed 19 of the 24 reported association types. In the different association types observed in Sistan Baluchistan province, number 14 (MAD20-K1-K1-MAD20) and number 17 (MAD-20-K1-K1-MAD20) were seen in the highest frequency (about 13%). The Vietnam study also had number 17 as the highest frequency (42%) but number 14 was only 12% of total associations observed.
Predominance of MAD20 type alleles have also been reported in Africa as early as 1991 by Conway et al. (1991) who used monoclonal antibodies recognizing polymorphic epitopes of MSP1 have shown the prevalence of MAD20 type paraistes in bock 6/18. As can be seen in our histogram, types 7, 8, 9,12 and 19 were not detected at all, in the Vietnam study too, these were represented poorly as 1, 0, 1, 1 and 6% of total frequencies observed.
The number of samples in this preliminary study were statistically significant though many more would be required to reach a final conclusion as to the reason for this variation including samples from Kerman province and Bandar Abbas, the two southern states from which P.falciparum is reported. This would require a much longer period of time as malaria is fortunately following a downward curve in our country.
ACKNOWLEDGMENTS
This study was supported by finance from Pasteur Institute of Iran, Tehran and National Center for Scientific Research, Tehran, Iran. I wish to thank Dr. Marcello U. Ferreira for his kind support for this study.
My acknowledgment to my colleagues, Ms. Fatemeh Mirkhani, Ms. Marzieh Tameemi and Ms. Arpi Hovanisian for their practical assistance.
REFERENCES
- Anderson, R.M., C.A. Dounley and S. Gupta, 1997. Vaccine design, evaluation and community based use for antigenically variable infectious agents. Lancet, 350: 1466-1470.
Direct Link - Cooper, J.A., L.T. Cooper and J.A. Saul, 1992. Mapping of the region predominantly recognized by antibodies to Plasmodium faciparum MSA1. Mol. Biochem. Parasitol., 51: 301-312.
Direct Link - Cowman, D., F. Alan, L.B. Deborah, H. Juli and E.M. Kerry et al., 2000. Functional analysis of proteins involved in P. falciparum merozoite invasion of RBCs. FEBS Lett., 476: 84-88.
Direct Link - Daubersies, P., S. Sallenave, S. Maque, J.F. Trape and H. Contamin et al., 1996. Rapid turnover of P. falciparum population in symptomatic individuals living in high transmission areas. Am. J. Trop. Med. Hyg., 54: 18-26.
Direct Link - Ferreira, M.U., W.L. Ribeiro, A.P. Tonon, F. Kawamoto and S.M. Rich, 2003. Sequence diversity and evolution of the malaria vaccine candidate merozoite surface protien -1 (MSP- 1). Gene, 304: 65-75.
Direct Link - Girard, P.H., 1988. Cerebellar syndrome in mild Plasmodium falciparum malaria. Trans. R. Soc. Trop. Med. Hyg., 82: 204-210.
Direct Link - Kaneko, O., M. Kimura, T. Kawamoto, M.U. Ferreira and K. Tanabe, 1997. Plasmodium falciparum: Allelic variation in the merozoite surface protein 1 gene in wild isolates from South Vietnam. Exp. Parasitol., 86: 45-57.
Direct Link - Da Silveira, L.A., M.L. Dorta, A.S.E. Kimura, A.M. Katzin and F. Kawamoto et al., 1999. Allelic diversity and antibody recognition of P. falciparum merozoite surface protein 1 during hypoendemic malaria transmission in the Barzilian Amazon region. Inf. Immun., 67: 5906-5916.
Direct Link - Miller, L.H., T. Robers, M. Shabuddin and T.F. McCutehn, 1993. Analysis of sequence diversity in the Plasmodium falciparum merozoite surface protein-1 (MSP-1). Mol. Biochem. Parasitol., 59: 1-14.
Direct Link - Peterson, M.G., R.L. Coppel, M.B. Moloneyu and D.J. Kemp, 1988. Third form of the precursor to the major merozoite surface antigen of Plasmodium falciparum. Mol. Cellular. Biol., 8: 2664-2667.
Direct Link - Pirson, P.J. and M.E. Perkins, 1985. Characterization with monoclonal antibodies of a surface antigen of P. falciparum merozoites. J. Immunol., 134: 1946-1951.
Direct Link - Riley, E.M., 1996. Thev role of MHC and non MHC associated genes in determining the human immune response to malaria antigens. Parasitology, 112: 39-51.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Tanabe, K., M. Mackay, M. Goman and J. Scaife, 1987. Allelic dimorphism in a surface antigen gene of the malaria parasite Plasmodium falciparum. J. Mol. Biol., 195: 273-287.
Direct Link - Tanabe, K., K. Murakami and S. Doi, 1989. Plasmodium falciparum Dimorphism of the p190 alleles. Exp. Parasitol., 68: 470-473.
Direct Link