
S.M. R. Ehteshami
Tarbiat Modares University, Tehran, Iran
M. Aghaalikhani
Department of Agronomy, Faculty of Agriculture, Tarbiat Modares University, P.O. Box 14115-336, Tehran, Iran
K. Khavazi
Soil and Water Research Institute, Tehran, Iran
M. R. Chaichi
Department of Agronomy, Tehran University, Karaj, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 20 | Page No.: 3585-3591







ABSTRACT
The effect of seed inoculation by phosphate solubilizing microorganisms on growth, yield and nutrient uptake of maize (Zea mays L. SC. 704) was studied in a field experiment. Positive effect on plant growth, nutrient uptake, grain yield and yield components in maize plants was recorded in the treatment receiving mixed inoculum of Glomus intraradices (AM) and Pseudomonas fluorescens (Pf). Co-inoculation treatment significantly increased grain yield, yield components, harvest index, grain N and P, soil available P, root colonization percentage and crop WUE under water deficit stress. In some of investigated characteristics under well-watered conditions, chemical fertilizer treatment was higher than double inoculated treatments, but this difference was not significant. Seed inoculation only with AM positively affected the measured parameters as amount as co-inoculated treatments. According to the results showed in contrast to the inoculated treatments with AM+Pf and AM, the application of alone Pf caused a comparatively poor response. Therefore, this microorganism needs to a complement for its activity in soil. All of measured parameters in inoculated treatments were higher than uninoculated treatments under water deficit stress conditions. Furthermore, the investigated characteristics of co-inoculated plants under severe water deficit stress conditions were significantly lower than co-inoculated plants under well-watered and moderate-stressed conditions. Therefore it could be stated, these microorganisms need more time to fix and establishing themselves in soil. The present finding showed that phosphate-solubilizing microorganisms can interact positively in promoting plant growth as well as P uptake of maize plants, leading to plant tolerance improving under water deficit stress conditions.
PDF Abstract XML References Citation
How to cite this article
S.M. R. Ehteshami, M. Aghaalikhani, K. Khavazi and M. R. Chaichi, 2007. Effect of Phosphate Solubilizing Microorganisms on Quantitative and Qualitative Characteristics of Maize (Zea mays L.) Under Water Deficit Stress. Pakistan Journal of Biological Sciences, 10: 3585-3591.
DOI: 10.3923/pjbs.2007.3585.3591
URL: https://scialert.net/abstract/?doi=pjbs.2007.3585.3591
DOI: 10.3923/pjbs.2007.3585.3591
URL: https://scialert.net/abstract/?doi=pjbs.2007.3585.3591
INTRODUCTION
Phosphorus as an essential mineral nutrient for plant growth and development is the world’s second highest chemical input in agriculture. Soluble P is often the limiting mineral nutrient for biomass production in agricultural ecosystems as well (Hameeda et al., 2006). Plants utilize fewer amounts of phosphate fertilizers that are applied and the rest is rapidly converted into insoluble complexes in the soil. So this phenomenon encourages farmers to frequent application of phosphate fertilizers. Modern agriculture is severely modifying and polluting the natural environment, due to the widespread application of chemical fertilizers, herbicides and pesticides. Therefore, thinking about valid alternative for chemical fertilizers is too necessary.
Microorganisms are important in agriculture because of it's role in promoting the circulation of plant nutrients and reduce the need for chemical fertilizers as much as possible (Cakmakci et al., 2006). Those microorganisms which dissolve poorly soluble CaP are termed as Mineral Phosphate Solubilizers (MPS). Phosphate solubilizing microorganisms (PSM) convert these insoluble phosphates into soluble forms through the process of acidification, chelation, exchange reactions and production of gluconic acid (Chung et al., 2005). Some of these microorganisms that colonize plant roots and promote growth when inoculate to seeds, roots or tubers have been termed plant-growth-promoting rhizobacteria (PGPR), because their application is often associated with increased rates of plant growth. Efforts to enhance the beneficial effects of PGPR by supplemental application to crops or soil were first reported in the 1950s from studies in the former Soviet Union and later in western countries (Sharma, 2002). In general, it is believed that PGPR are more effective in promoting plant growth under limited supply of nutrients. Similar results were reported in some of the previous studies showing that inoculation was found to affect plant growth and development, plant dry weight and grain yield of plants (Dobbelaere et al., 2002). Many PGPR belong to the fluorescent pseudomonas, including P. fluorescens that can influence plant growth directly or indirectly by releasing mineral nutrients and phytohormones (Rincon et al., 2005). The previous observations have indicated that the growth and yield of different crops increased with the inoculation of P. fluorescens (Shaharoona et al., 2006). The rhizoplane and surrounding soil rhizosphere are colonized or otherwise occupied by a wide range of microorganisms. Of the various microorganisms present, Vesicular Arbuscular Mycorrhiza (VAM), the most widespread and agronomically important type of plant symbiont, has a great value in promoting uptake of phosphorus, nitrogen; micronutrients and water lead to increase growth and yield of several crops (Wang et al., 2006). It was found that VAM fungi interact synergistically with other microorganisms such as phosphate-solubilizing bacteria and plant growth promoting microorganisms to enhance plant growth and survival, also a significant effect of bacteria producing plant growth regulators on mycorrhizal development has been reported (Azcon, 1989).
Seasonal drought and water deficit stress are considered as the most important abiotic stress that limits crop production in arid and semi-arid areas (Kramer and Boyer, 1997). Arbuscular mycorrhizal fungal symbiosis is widely believed to protect host plants from detrimental effects of drought. It has been shown that AM association enhances P uptake by plant roots under drought. Improved P nutrition by AM fungi during the periods of water deficit has been postulated as a primary mechanism for enhancing host plant drought tolerance (Subramanian et al., 2006).
The main purpose of this study, is investigating the effects of Glomus intraradices and Pseudomonas fluorescens strain 93 inoculations on quantitative and qualitative characteristics of maize (Zea mays L. SC. 704). We have tried to understand their ability to make a successful symbiosis with the crop and finding its ecophysiological role in maize growth and development under water deficit stress conditions.
MATERIALS AND METHODS
Experimental site and design: This experiment was conducted at Meshkin-Abad experimental station of Soil and Water Researches Institute of Iran, during June-October 2005. The soil texture was loam and had 6.2 mg kg-1 available P, 135 mg kg-1 nitrogen; 170 mg kg-1 potassium and pH = 7.66. The field had been under fallow for 2 former years; hence the soil had a low indigenous VAM and phosphate-solubilizing bacteria populations.
The experiment design consisted of four randomized complete blocks in a split-factorial arrangement having 15 treatments in each block. The main-plots regarding to simulating the water deficit stress levels consisted of three water regimes, which was achieved by scheduling cumulative pan evaporation in mm. The irrigations were scheduled for various treatments, when the cumulative pan evaporation readings reached at 70, 100 and 130 mm. The sub-plots included of the application of microbial inoculants [Glomus intraradices; Pseudomonas fluorescens strain 93; Glomus intraradices + Pseudomonas fluorescens strain 93 (50-50%)] and not [triple super phosphate fertilizer; without fertilizer (control)].
The experimental area was ploughed, rotavated and divided into 4 blocks separated by a 2 m wide path. According to soil analysis test all treatments received recommended amounts of Urea and K2O fertilizers that drilled in sowing time. Seventy five percent of the urea fertilizer was applied in two equal splits at 3-leaves and 10-leaves stages. Maize (Zea mays L. SC.704) inoculated seeds were hand sown at 20 cm spacing in rows 75 cm apart. All plots were hand-weeded and irrigated uniformly till maize 8-leaves stage. In this stage, plants were exposed to intensities of water deficit stress (severe, moderate and no water deficit), the depth of irrigation was maintained consistent at all of treatments and the quantity of irrigation water delivered to each plot was measured using a water contour.
Bio-inoculants characterization and seed treatment: P. fluorescens was in the collection and used strain (strain 93) was isolated from the rhizosphere of wheat (Triticum aestivum L.) by Soil Biology Laboratory in Soil and Water Researches Institute of Iran. The inoculum of Glomus intraradices consisted of AM propagules. This inoculum was uniformly mixed into the apatite that was prepared by Mycorrhiza Laboratory in mentioned Institute. Maize seeds were inoculated and then placed in the furrow.
Plant data: The last four expanded leaves of maize were sampled for plant tissue’s P analysis before tasselling. At harvest, the central 4 m2 area of each plot was harvested and grain yield, yield components and harvest index were recorded. Shoots were oven dried at 75°C until a constant weight was obtained. Water Use Efficiency (WUE) calculated by dividing the total grain yield (kg ha-1) to the quantity of consumed water (mm).
Soil and plant analysis: The plant tissue’s samples were washed thoroughly, dried at 75°C, weighed and digested in di-acid mixture in sealed chamber. The digested samples were diluted to 50 mL with distilled water. The phosphorus content was estimated using standard micro-kjeldahl and vanadomolybdo phosphoric acid yellow color methods, respectively. For nutrient analysis, the following procedure was applied: 1 g seed was digested in an acid mixture of HNO3 and HClO4 (9:4). The resulting ash was analysed for nutrient content. Total nitrogen was determined by Kjeldhal’s method. Phosphorus content was determined with the molybdenum-ascorbic acid colorimetric method. Soil P was extracted from 2 g moist soil with 40 mL 0.5M NaHCO3 at pH 8.5.
The percentage of root colonization: Roots were located in a solution of 50% ethanol and 50% distilled water. A portion of the root sample was thawed and after clearing the roots with KOH and staining the roots in 0.05% trypan blue, percentage of root segments colonized by G. intraradices was estimated by the line-intersect method (Kormanik and McGraw, 1982).
Statistical analysis: The statistical analysis of data was carried out by conducting ANOVA. Comparison of the means was made using Least Significant Difference (LSD). All the data are presented as actual (non-transformed) means. Percentage values were arc sin-transformed before statistical analysis.
RESULTS
Grain yield and yield components: The grain yield of maize decreased (p≤0.05) under varying intensities of water defecit stress. Conversely, Glomus intraradices (AM) and Pseudomonas fluorescens (Pf) bio-inoculants enhanced the maize grain production regardless of intensities of water deficit stress, but this response was more pronounced under moderate water deficit conditions than in sever water deficit treatments. The dual inoculation showed significant difference in grain yield of maize comparing to control (uninoculated). Inoculation with AM + Pf and AM under well-watered conditions increased the grain yield compared with other treatments, but had a lesser effect than P fertilizer treatment. However, this difference was not statistically significant. Inoculation with AM enhanced the grain yield under different water deficit stress conditions in compared with P fertilizer too. The highest increase in 100-grain weight, of row number per ear and grain number per row was recorded in P fertilizer treatment, but this difference was not statistically significant (p≤0.05). In the presence of AM+Pf and AM treatments, yield components (expect row number per ear) enhanced significantly in compared with P fertilizer and control, when plants were exposed to water deficit conditions (Table 1).
The water deficit treatment significantly (p≤0.05) decreased yield components of control (uninoculated) and P fertilizer treatments. While water deficit stressed AM + Pf and AM treatments were comparable to well-watered and fertilizered plants. Single inoculation with Pf did not cause plants to differ significantly in grain production and yield components. Harvest index was also higher in plots supplied with P fertilizer than in plots with co-inoculation, but this difference was not significant. The water deficit treatments decreased HI for inoculated and non-inoculated plants, but HI values were higher in co-inoculants and treatments than P fertilizer and control plants. The HI values for water deficit stressed AM+Pf and AM plants were comparable to well-watered treatments (Table 1).
Minerals concentration: Phosphorus concentration in plant tissue were increased in plants by inoculation with AM+Pf and AM under well-watered and water deficit stress condition. The inoculation with AM+Pf significantly (p≤0.05) increased the P contents of maize plants and had higher than control and P fertilizer plants under varying intensities of water deficit stress. P uptake by plant was also more with the application AM under moderate stress in our study. The concentration of Phosphore in control plants was significantly (p≤0.05) lower than that in plants grown under treatments AM+Pf and AM (Table 2).
The dual inoculation of Pf and AM resulted in a significant increase of grain P and N concentration (Table 2). The P and N concentration in treatments of AM+Pf, AM and P fertilizer increased by 200, 32.47, 182.35, 24.11, 176.47 and 23.53%, respectively in comparison to the uninoculated control under water-welled conditions. The inoculation with AM had a more stimulating effect on the assimilation of P and N in comparison with alone bacterial inoculation. However, AM performed better than P fertilizer, but was less than AM + Pf in stimulating N and P uptake. The trend of grain P and N accumulation under different treatments of water deficit stress was similar to well-watered conditions. The P and N concentration of grain in plants under different treatments ranged from 0.132, 1.14 (control) to 0.51, 1.852 (dual inoculation with AM + Pf), respectively. The lowest P and N concentration of grain was detected in plants grown in uninoculated and unfertilised treatments.
| Table 1: | Maize grain yield and yield components for phosphate fertilizer (P), Glomus intraradices (AM), Pseudomonas fluorescens (Pf), co-inoculation (AM+Pf) and no fertilizer (Control) plants exposed to varying intensities of water deficit stress (70, 100 and 130 mm cumulative pan evaporation) |
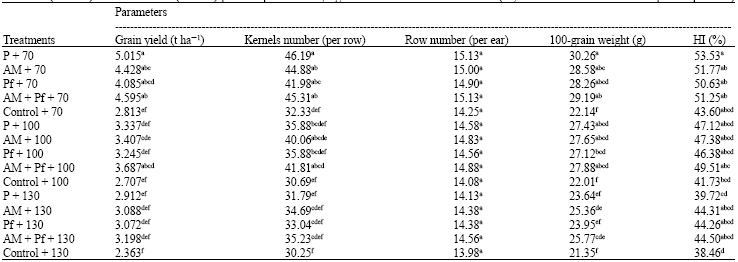 | |
| Means with different superscript letter(s) are significantly different at p≤0.05 according to LSD test | |
| Table 2: | Minerals concentration (%), in plant tissue and soil available P content (mg kg-1 soil) measured for phosphate fertilizer (P), Glomus intraradices (AM), Pseudomonas fluorescens (Pf), co-inoculation (AM+Pf) and no fertilizer (Control) plants exposed to varying intensities of water deficit stress (70, 100 and 130 mm cumulative pan evaporation) |
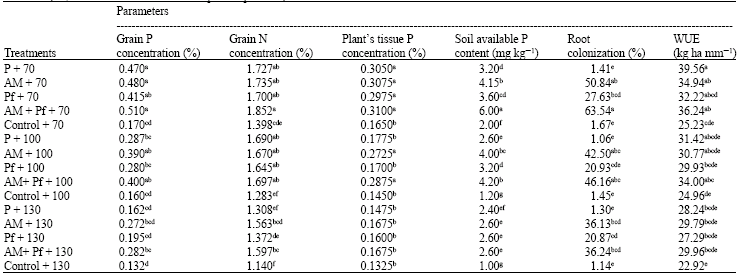 | |
| Means with different superscript letter(s) are significantly different at p≤0.05 according to LSD test | |
Treatment of AM+Pf and AM inoculation resulted in a significant increase in P and N uptake to different degrees of water deficit stress when compared with the control. Although inoculated plants with Pf showed unexpectedly low grain P and nitrogen content (Table 2).
The highest soil available P was associated with plants grown under AM+Pf co-inoculation treatment. This value was significantly (p≤0.05) higher than that of plants grown under all other treatments under similar conditions. Present study showed that inoculated maize plants by AM + Pf are much more efficient in taking up soil P than non-inoculated plants. All inoculation treatments significantly improved available P content of soil. The available P in soil under AM + Pf and AM were greater than single-inoculation treatments too (Table 2).
Root colonization: The percentage of root colonization was significantly higher in the treatments containing co-inoculants and AM than the untreated control and P fertilizer treatments. The highest increase in the percentage of root colonization was recorded by co-inoculated treatments (63.54%). Present results showed water deficit stress decreased the percentage of root colonization. The mycorrhizal colonization in control and P fertilized plants remained less than 3% regardless of irrigation treatments (Table 2).
Water use efficiency: The water use efficiency (WUE) of inoculated treatments was higher than non-inoculated treatments in moderate and severe intensities of water deficit stress. In spite of no significant difference between inoculation treatments, inoculation with AM+Pf and AM under well-watered conditions increased WUE compared with other treatments, but had a lesser effect than P fertilizer treatment.
In contrast to the inoculated treatments with AM+Pf and alone AM, the application of alone Pf cause a comparatively poor response in all of measured parameters (Table 2).
DISCUSSION
The inoculation by Glomus intraradices (AM) and Pseudomonas fluorescens (Pf) improved growth, water status, yield, yield components, grain P and N and soil P, when exposed to varying intensities of water deficit stress. In this study, the beneficial effect of AM+Pf inoculation in terms of increased grain production was more revealed under water deficit conditions. The improved nutritional status and relative water content caused by dual inoculation would have alleviated water deficit impacts and promoted maize grain yield and its yield component under varying intensities of water deficit stress compare to uninoculation treatments. These induced changes by AM+Pf indicate a possible involvement of plant hormones in the effects of inoculants on plants. It is reported that fluorescent pseudomonas produce numerous metabolites such as plant growth regulators and vitamins, which affect the growth of plants and microorganisms present in soil (Prikryl et al., 1985). All these products are small molecules and can pass through the membrane during ultrafiltration. These microorganisms could further influence the behaviour of AM propagules through the production of bioactive compounds (Vosatka and Gryndler, 1999). Microorganisms producing phytohormones cohabit in the rhizosphere with AM fungi and could play an additive role in the plant-fungus relation. But this possible interactive effect needs to be proved. As previously reported Leinhos (1994) these results indicate the existence of Functional compatibilities between saprotrophic and symbiotic microorganisms. The plant response may be diagnostic for the success of the association which is dependent on the AM fungi involved. It is known that the presence of tryptophan, as a precursor of auxin, stimulates bacterial IAA production. Therefore, the production of IAA, in root exudates, by the still-living cells of P. fluorescens can result from utilisation of tryptophan released by dying cells. In contrast, amino acids like asparagine, alanine and lysine, known to be present in root maize exudates, can stimulate the activity of enzyme like tryptophan aminotransferase (Martens and Frankenberger, 1994). Moreover, sugars, present in exudates, could be used as C-source by bacteria; Leinhos (1994) has shown that these compounds had an effect not only on plant growth but also on auxin production by Pseudomonas sp. The bacterium-fungus association increases this effect.
The P content of co-inoculated maize plants was consistently higher than uninoculated plants regardless of intensities of water deficit stress. A close relationship between P content and water deficit tolerance has been reported earlier (Subramanian et al., 1997). Present field study suggests that AM + Pf inoculation improves water deficit tolerance of maize plants as a secondary consequence of enhanced nutritional status of the host plant, especially P. It is known that vesicular-arbuscular mycorrhizal (VAM) fungi and PGPR can increase P uptake by plants through their phosphatase activity, through a better exploration of the soil by hyphae and through the uptake of fixed soil P, which is unavailable to plant roots. Our findings emphasize the dependence of maize plants on root colonization for P absorption. As, our data agree with the findings of Sylvia et al. (1993), who reported that mycorrhizal colonization with Glomus fasciculatum improved the drought tolerance of field-grown maize plants as a result of enhanced P status under varying intensities of drought stress.
A combined bio-inoculation of PGPR strain and AMF had a significant effect on grain quality. For instance, the phosphorus concentration of grain increased in the bio-inoculated plants. Interestingly, a synergistic effect on the level of N concentration was observed between AMF and PGPR when they were co-inoculated too.
The available P was greater in soil under co-inoculation treatments. The experimental data agree with the findings of others that inoculants increase soil available P (Zaidi and Khan, 2005). The poor availability of P in the soil samples could be due to uptake of P by the crop, or to chemical fixation of P.
Mycorrhizal colonization of maize roots was increased in the treatments inoculated with AM and Pf. The greater effectiveness of this co-inoculant might be related to its high root colonization ability and chitinase activity in addition to ACC-deaminase activity (Shaharoona et al., 2006). Others have suggested that the host plant drought tolerance resulting from root colonization may be explained by a greater absorption root surface area or densely proliferated root growth or hydraulic differences between root systems (Subramanian et al., 2006). Therefore, improving the area of soil in contact with the plant root increase the absorptive surface area of the mycorrhizal root system and extend the P-depletion zone away from the root surface.
Since inoculated treatments AM+Pf and alone AM consistently increased grain yield under varying intensities of water deficit stress, WUE of these plants were much higher than well-watered control plants. The experimental data agree with the findings of others that support mycorrhizae-aided host plant drought tolerance and associated yield increase under water deficit conditions (Subramanian et al., 2006).
The inoculation of alone Pf caused a comparatively poor response in all of measured parameters, which could possibly be due to the poor ability of this microorganism to solubilize inorganic P already present in soil alonely. Therefore, our findings showed this microorganism needs to a complement for its activity in soil. Previous studies involving plants inoculated with PSMs showed growth enhancement and increased P contents but large variations were found (Zaidi and Khan, 2005).
Present results showed maize plants did not respond to dual inoculation in severe water deficit stress conditions. It is reported water uptake by entire root systems is determined by the amount of roots, their distribution in soil and the rate of water absorption per unit root (Auge, 2001). Regarding to our results, the percentage of root colonization decreased when water deficit stress increased of moderate to severe stress conditions. Therefore, it is seems water and P uptake rates per unit root length (specific water and P uptake rates) affect by root colonization. On the other hand, it is recognized both symbiosis and species richness of VAM fungi are reportedly lower in arid climates than in other ecosystems and decrease as aridity increases (Auge, 2001).
CONCLUSIONS
In summary, AMF and PGPR bio-inoculants improved drought tolerance of field grown maize plants as a consequence of enhanced nutritional status especially P. AMF and PGPR bio-inoculants response was more pronounced under water deficit stress than well-watered conditions. The data revealed that mentioned bio-inoculants enables the host plant to withstand varying of water deficit stress under field conditions. Indeed, these bio-inoculants were adapted to their environment in terms of soil characteristics, plant genotype and climate. Meanwhile, this research must be proactive and the field trials must be established across a broad range of soil and environmental conditions and must be conducted within the context of current or future farming practices.
REFERENCES
- Auge, R.M., 2001. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza, 11: 3-42.
CrossRefDirect Link - Azcon, R., 1989. Selective interaction between free-living rhizosphere bacteria and vesiculararbuscular mycorrhizal fungi. Soil Biol. Biochem., 21: 639-644.
CrossRefDirect Link - Cakmakci, R., F. Donme, A. Aydin and F. Sahin, 2006. Growth promotion of plants by plant growth promoting rhizobacteria under greenhouse and two different field soil conditions. Soil Biol. Biochem., 38: 1482-1487.
Direct Link - Chung, H., M. Park, M. Madhaiyan, S. Seshadri, J. Song, H. Cho and T. Sa, 2005. Isolation and characterization of phosphate solubilizing bacteria from the rhizosphere of crop plants of Korea. Soil Biol. Biochem., 37: 1970-1974.
CrossRefDirect Link - Dobbelaere, S., A. Croonenborghs, A. Thys, D. Ptacek, Y. Okon and J. Vanderleyden, 2002. Effect of inoculation with wild type Azospirillum brasilense and A. irakense strains on development and nitrogen uptake of spring wheat and grain maize. Biol. Fertil. Soils, 36: 284-297.
CrossRefDirect Link - Leinhos, V., 1994. Effects of pH and glucose on auxin production by phosphate-solubilizing rhizobacteria in vitro. Microbiol. Res., 149: 135-138.
Direct Link - Martens, D.A. and W.T. Frankenberger Jr., 1994. Assimilation of exogenous 2′-14C-indole-3-acetic acid and 3′-14C-tryptophan exposed to the roots of three wheat varieties. Plant Soil, 166: 281-290.
CrossRefDirect Link - Rincon, A., B. Ruiz-Diez, S. Garcia-Fraile, J.A.L. Garcia, M. Fernandez-Pascual, J.J. Pueyo and M.R. deFelipe, 2005. Colonization of Pinus halepensis roots by Pseudomonas fluorescens and interaction with the ectomycorrhizal fungus Suillus granulatus. FEMS Microbiol. Ecol., 51: 303-311.
Direct Link - Shaharoona, B., M. Arshad, Z.A. Zahir and A. Khalid, 2006. Performance of Pseudomonas spp. containing ACC-deaminase for improving growth and yield of maize (Zea mays L.) in the presence of nitrogenous fertilizer. Soil Biol. Biochem., 38: 2971-2975.
CrossRefDirect Link - Subramanian, K.S., P. Santhanakrishnan and P. Balasubramanian, 2006. Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Sci. Hortic., 107: 245-253.
CrossRefDirect Link - Sylvia, D.M., L.C. Hammond, J.M. Bennett, J.H. Haas and S.B. Linda, 1993. Field response of maize to a VAM fungus and water management. Agron. J., 85: 193-198.
Direct Link - Vosatka, V. and M. Gryndler, 1999. Treatment with culture fractions from Pseudomonas putida modifies the development of Glomus fistulosum mycorrhiza and the response of potato and maize plants to inoculation. Applied Soil Ecol., 11: 245-251.
CrossRefDirect Link - Wang, F., X. Lin, R. Yin and L. Wu, 2006. Effects of arbuscular mycorrhizal inoculation on the growth of Elsholtzia splendens and Zea mays L. and the activities of phosphatase and urease in a multi-metal-contaminated soil under unsterilized conditions. Applied Soil Ecol., 31: 110-119.
Direct Link - Zaidi, A. and M.S. Khan, 2005. Interactive effect of rhizotrophic microorganisms on growth, yield and phosphorus uptake of wheat. J. Plant Nutr., 28: 2079-2092.
CrossRefDirect Link