
A. Karimi
Department of Animal Science, University of Razi, Iran
H. Karami
Department of Animal Science, University of Razi, Iran
M. M. Moini
Department of Animal Science, University of Razi, Iran
A.A. Ahmadi Sefat
Department of Animal Science, University of Razi, Iran
E. Haghvirdilu
Department of Animal Science, University of Razi, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 20 | Page No.: 3557-3563







ABSTRACT
In order to determine different protocols of synchronization and their efficiency on pregnancy rate after fixed-timed AI (TAI), 120 dairy Holstein cows (n = 120) were assigned randomly to six groups: 1) two injections of Prostaglandin F2α (PGf2α) with 12 days apart as a control group, 2) two injections of Gonadotropin Releasing hormone (GnRH) with 9 days apart and an injection of PGf2α at day 7, 3) injection of GnRH and PGf2α with 7 days apart, then single injection of Estradiol Benzoate (EB) after 48 h, 4) injection of progesterone (P4) in conjugation with EB then after 7 days PGf2α injection and after 48 h an injection of GnRH were done, 5) as group 4 but EB was used instead of GnRH, 6)injections of PGf2α and EB conjugated with Human Chronic Gonadotrophin (hCG) with 12 h apart. Animals in group 1 (control), groups 2-5 and group 6 were inseminated after 72, 20 and 36 h, respectively. Serum P4 concentration of group 4 (4.43± 1.50 ng mL-1) was higher than control group (2.34±1.36 ng mL-1) at day 5 after insemination (p<0.05); P4 concentrations of groups 3 and 4 have significant differences with control group (2.69±2.64 and 2.56±1.40 versus 0.81±0.41 ng mL-1, respectively, p<0.05) at a day after second injection and groups 4 and 5 were in higher level of P4 concentration than control group at insemination time (3.14±1.9 and 2.89±1.8 versus 0.45±0.19 ng mL-1 respectively, p<0.05). Pregnancy rate were 0, 50, 45, 10, 30 and 45% for group 1 (control) through 6, respectively.
PDF Abstract XML References Citation
How to cite this article
A. Karimi, H. Karami, M. M. Moini, A.A. Ahmadi Sefat and E. Haghvirdilu, 2007. Comparison Between Different Protocols of Synchronization and Their Efficiency on Pregnancy Rate of Dairy Cattle. Pakistan Journal of Biological Sciences, 10: 3557-3563.
DOI: 10.3923/pjbs.2007.3557.3563
URL: https://scialert.net/abstract/?doi=pjbs.2007.3557.3563
DOI: 10.3923/pjbs.2007.3557.3563
URL: https://scialert.net/abstract/?doi=pjbs.2007.3557.3563
INTRODUCTION
Synchronisation of oestrus in cattle has been used to facilitate the use of Artificial Insemination (AI) and to minimize the time needed to detect oestrus in cattle (Cavalieri et al., 2005). Precise control of the bovine estrous cycle requires control of follicular waves and Corpus Luteum (CL) lifespan (Ahuja et al., 2005; Patterson et al., 2000). Oestrous synchronization protocols typically employ one of two strategies: 1) administration of a luteolytic agent such as prostaglandin f2α or 2) application of a native or synthetic progestagen to delay the time of estrus following natural or induced luteolysis which may extend the length of the estrous cycle, by CIDR etc (Macmillan and Peterson, 1993; Martinez et al., 2002; Kim et al., 2003; Mialot et al., 2003). Administration of PGF2α analogues induces rapid, premature luteal regression in females with a functional Corpus Luteum (CL) (Kristula et al., 1992), followed by spontaneous ovulation (Khanum et al., 2006; Ferguson and Galligan 1993; Folman et al., 1990). The current and future direction of estrous synchronization is to focus on combining traditional methods of controlling cycle length with the manipulation of follicular development in order to program or select the ovulatory follicle. The immediate goal of controlling both CL function and follicular development is to devise a treatment that will synchronize estrus more precisely and to control the time of ovulation more exactly to allow a single timed insemination without the need for detection of behavioral estrus. The ultimate goal may be to improve the fertility of lactating cows and heifers over that of untreated animals exhibiting a spontaneous estrus. The mode of action of prostaglandins (PG) in the control of ovulation is to induce a premature luteolysis and consequent fall in circulating progesterone concentrations by removal of negative feedback inhibition on the hypothalamus and pituitary, the fall in progesterone concentrations allows a sequence of hormonal and ovarian events that culminate in oestrus and ovulation (Peters, 2005). Recent studies have incorporated GnRH into a synchronization protocol (Are´chiga et al., 1998; Burke et al., 1996; LeBlanc et al., 1998; Pursley et al., 1997a; 1995; Schmitt et al., 1996; Stevenson et al., 1996; Twagiramungu et al., 1995) that involves a series of hormone administrations. Ovsynch [synchronization of ovulation (GnRH, day 0; PGf2a day 7; GnRH, day 9; then timed artificial insemination (AI) after second GnRH)] is a management tool that uses GnRH and PGf2a to synchronize ovulation, thus allowing control of first and subsequent timed AI in dairy cows (Pursley et al., 1997a) and has been shown Pregnancy Rate (PR) to a timed AI following treatment with Ovsynch is similar to AI after observed estrus (Stevenson et al., 1999; Britt and Gaska, 1998; De la Sota et al., 1998; Burke et al., 1996; Pursley et al., 1997a; Momcilovic et al., 1998; Pursley et al., 1997b).
Preparations of GnRH, Human Chorionic Gonadotropin (hCG), LH and P4 are frequently used to treat synchronization of ovulation (Trigg, 2004; Todoroki et al., 2001, Tebble et al., 2001; Garverick 1997; Osawa et al., 1995; Peter 1998; Thatcher et al., 1993; Calder et al., 1999) but treatment outcomes are highly variable. Recent reports indicate that ovsynch, followed by timed AI performed 16-20 h after the 2nd GnRH treatment yields pregnancy rates of about 25% in cows with occurrence of synchronization of ovulation (Bartolome et al., 2000; Fricke and Wiltbank, 1999).
Pharmaceutical control of the reproductive cycle (PCRc) has some advantages and disadvantages. PCRc can remove the estrous detection (Cavalieri et al., 2005), also there may be some advantages in shortening the calving to conception interval (Ryan et al., 1999), therefore calving interval is reduced and etc. currently the major disadvantage of PCRc is that reproductive performance is highly variable and results vary considerably between herds and between years in the same herd (Padula and MacMillan, 2002).
Hence we conduct an experiment with six different hormone programs in order to determine the best program for shortening the calving to conception interval and pregnancy rate of each program following after timed AI and their hormonal profile position. Here, we particularly used a program for rapid ovulation induction in comparison with other programs of ovulation synchronization.
MATERIALS AND METHODS
Location of farm and date of experiment: This study was conducted on a 1600-Holstein dairy cow in Kermanshah province of Iran. The farm is located in Ravansar region between Latitudes 34° N and 35° N and between Longitudes 46° E and 47° E. The climate of the area is characterized by an average annul of 14.9°C and 537.9 mm mean rainfall per year. The study period lasted almost 3 months from January 2006 to April 2006.
Treatments: One hundred twenty dairy cows (BW = 645±25.245 Kg, Number of parity = 2.8±1.46, Days after Postpartum = 34.6±3.1 and Milk yield = 7358.23±1280.79) were grouped in six groups (Fig. 1):
| • | In control group each cows received two injections of 500 mg of the prostaglandin F2α (PGf2α) analogue, cloprostenol sodium (manufactured by Nasr pharmaceutical company, Fariman, Iran), with 12 days apart. |
| • | In this group 500 mg two injections of Gonadorelin (an analog of GnRH/manufactured by Aburaihan Pharmaceutical Co. Veterinary Division Tehran, Iran) were performed with 9 days apart and also 500 mg PGf2α was injected in day 7. |
| • | At first, all cows were injected by 500 mg GnRH and 7 days later were injected 500 mg PGf2α and then, 2 mg Estradiol Benzoate (EB) (manufactured by Aburaihan pharmaceutical Co. veterinary division Tehran, Iran) was used after 48 h. |
| • | At first, there was an injection of 2 mg EB conjugated with progesterone (manufactured by Aburaihan Pharmaceutical Co, Tehran, Iran) and 7 days later, an injection of 500 mg PGf2α was used and 48 h later 500 mg GnRH was injected. |
| • | Here animals received 2 mg EB conjugated with 100 mg progesterone; after 7 days 500 mg PGf2a was injected then 48 h later 2 mg EB was used. |
| • | In this group 500 mg PG was used per cow and after 24 h animals received 1 mg EB conjugated with 250 IU of Human Chorionic Gonadotropin (hCG) (manufactured by Daroupakhsh pharmaceutical Co. Tehran, Iran). |
Estrus detection, insemination and pregnancy determination: In this report, estrous rate have been shown as the percentage of cows in a group that showed estrous activity.
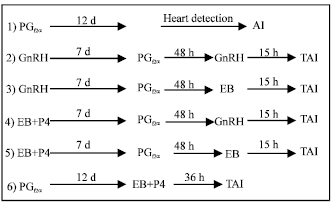 | |
| Fig. 1: | Schematic representation of synchronization protocols used in each experiment. (PGf2a = prostaglandin F2α, GnRH = Gonadotropin Releasing hormone, EB = Estradiol Benzoate, P4 = progesterone, hCG = Human Chorionic Gonadotropin, TAI = Timed Artificial Insemination, d = Day, h = Hour) |
Detection of estrous was carried out by heatman as a routine daily activity for monitoring of standing estrous.
Also Artificial Insemination (AI) was carried out as a routine activity via frozen-tawed semen of a same bull.
All cows of group1(control) were inseminated after 72 h but other cows of group 2-5 and cows of group 6 were inseminated after 20 h and after 36 h, respectively. Then animals were examined for pregnancy determination by palpation, 40 days post insemination and pregnancy rate was defined as percentage of cows conceived after AI in a group.
Blood collection and progesterone assay: Blood sampling for determination of serum P4 concentration in group 1 (as control group) through 5 was performed before the first injection (day = 0), day 5 (2 days before second injection), day 8 (a day after second injection) and at insemination time, exactly before AI. But in group 6 that was short-length induction of ovulation, samples were collected before first injection, a day after second injection and exactly before AI. (Because this treatment was short-length induction and there was no sample in 2 days before the second injection).
Blood samples were taken from coccygeal vein. Then blood samples stored stably and allowed to clot at 4°C then centrifuged 2000 g for 10 min and separated serum stored at -20°C until progesterone assay. Serum P4 concentrations were determined using an extract single antibody Radioimmunoassay (RIA).
Data and statistical analysis: The parity number, days in postpartum period and milk yield were the factors that used for grouping of cows. Analysis of serum P4 concentrations between groups was performed via ANOVA procedure in the same time of sampling. The data analysis have expressed as mean±SD that carried out in SPSS program at a p-value less than 5% (p<0.05) as significant level using Duncan’s multiple range test (Duncan, 1955).
RESULTS
There were no significant differences between experimental animals in number of parity (2.8±1.46), days after postpartum (34.6±3.1) and milk yield (7358.23 ±1280.79) (p>0.05).
After RIA of progesterone, it was found out a cow of group4 was in luteal cystic disease, therefore it was removed from trial.
The average of plasma progesterone concentrations on the start day of trial was 2.43±2.16 ng mL-1 and there is no difference between experimental animals (p>0.05). There is a significant difference in serum P4 concentration of second blood sample of group 4 than control and group 2 and 3 (4.43± 1.50 versus 2.34±1.36, 2.9±1.69, 3.14±2.34 ng mL-1, respectively, p<0.05), (Table 1).
Group 3 and 4 had significant differences versus control group in serum p4 concentrations of third blood sampling (2.69±2.64 and 2.56±1.40 versus 0.81±0.41 ng mL-1, respectively, p<0.05), (Table 1).
Also, Duncan’s multiple range tests shows a higher level of serum p4 concentrations in fourth blood sampling, between group 4 and 5 (3.14±1.9 and 2.89±1.8 ng mL-1, respectively) versus other groups (p<0.05) (Table 1).
Number of cows exhibited estrous were 40, 45, 65, 35, 40 and 60% and pregnancy rate were 0, 50, 45, 10, 30 and 45% for group 1(control) through 6, respectively (Table 1).
| Table 1: | Serum P4 concentrations at various blood sampling, estrus rate and pregnancy rate of experimental groups |
 | |
Different letter(s) (a, b and c) in each blood samples show a significant difference between groups (p<0.05). 1. G = Experimental group 2. B.S = Blood Sample (B.S1: day 0, before the first injection; B.S2: 2 days before second injection; B.S3: In a day after second injection; B.S4: At insemination time, exactly before AI. *In group6 that was short-length induction of ovulation, samples were collected before first injection, a day after second injection and exactly before AI. (Because this treatment was short-length induction and there was no sample in 2 days before the second injection), **A cow of group4 was in luteal cystic disease | |
DISCUSSION
The mean P4 concentration was 2.24 ng mL-1 in control group before first injection. Mean P4 concentration was 2.34 ng mL-1 10 days after first injection. Using two injections of PGf2α will bring most females in to heat 2-5 days after the second injection (Khanum et al., 2006). The mean P4 concentration was 0.816 ng mL-1 one day after second injection of PGf2α. This reduction in P4 concentration shows the effect of PGf2α on CL regression. Theoretically, only 55% of cows have sensitive CL to prostaglandin (Taveren, 1992). Therefore, two injections of PGf2α at 11-14 days apart is necessary to bring all cows in synchronization programs (Burk et al., 1997). Although estrous synchronization with PGf2α is simple, but has different disadvantages such as variety in pregnancy rate in response to stage of estrous cycle at the time of PGf2α injection (Rayon et al., 1998).
Investigation of P4 concentration of second blood samples show presence of ovarian CL in the majority of the animals in group 2 and 3. Response to GnRH treatment is associated with growing phase of dominant follicles (Trimberger, 1948). Predominantly, ovulation is stimulated during the end of growing phase that there are sufficient LH receptors on granulose cells (Webb et al., 1999). Reduction in P4 concentration of third blood samples shows CL regression after PGf2α administration and a decrease of less than 1 ng mL-1 after second GnRH injection at AI time. Group 3 was similar to group 2 except that EB was used instead of GnRH which induced heat, LH surge and ovulation (Hobson and Hansel, 1972).
In group 4 mean P4 concentrations in the first and second sample were 2.72 and 4.43 ng mL-1, respectively. After PGf2α injection, P4 concentration decreased (2.56 ng mL-1) in third sample. Finally, after GnRH agonist injection, mean P4 concentration increased (3.14 versus 2.56 ng mL-1) in fourth samples before TAI. SerumP4 concentration of the second sample was higher than the first and third ones. These differences show the effect of PGf2α on CL regression. Administration of estradiol in the presence of luteal phase progesterone or an exogenous progesterone source has also been shown to induce follicle atresia and to suppress gonadotropin secretion (Stevenson et al., 1984; Michael et al., 2000). Treatment with a combination of EB and progestagen has considerable promise for causing follicle atresia and synchronizing follicular development for both estrous synchronization and superovulation (Thatcher et al., 2002). Therefore injection of combination of EB and progesterone caused follicular atresia and induction of a new follicular wave and this is the way that in which mean progesterone concentration arrived to 4.43 ng mL-1 in second sample.
Administration of GnRH 10, 30 or 50 h after PGf2α likewise reduced the number of cows responding to PGf2α. Administration of GnRH 52 h after PGF2α resulted in a better synchronization of ovulation than PGf2α alone, but conception rate was reduced if the GnRH injection was given 60 or 72 h after the PGf2α (Gilbert, 1997). In present study, GnRH injected 48 h after PGf2α treatment and similar to previous works, the estrous and pregnancy rate was low. Increase in P4 concentration after GnRH injection, probably is due to LH secretion and its positive effects on luteinization of granulosa cells.
In group 5 after joint injection of EB and progesterone, the P4 concentration decreased to 3.74 ng mL-1 in second sample versus first one and after PGf2α injection decreased to 1.61 ng mL-1 in third sample and finally after EB injection, increased in fourth one (2.89 ng mL-1). Similar to group 4 joint injection of EB and progesterone caused follicle atresia then new follicular wave induced and mean P4 concentration arrived to 3.74 ng mL-1 in second sample.
The interval from the PGf2α injection to onset of the heat varies depending on the stage of the cycle when PGf2α is administered. Therefore, the age of the CL and status of developing follicle determine time to the onset of the heat following PGf2α injection (Lopez, 2000, Macmillan and Peterson, 1993). Positive feedback loop between estradiol and LH is critical in enabling on ovarian follicle to mature and ovulate and facilitating expression of estrous coincident with ovulation (Schmitt et al., 1995). Therefore, injection of EB after PGf2α, in this group, caused LH surge and ovulation, therefore mean progesterone concentration before artificial insemination increased.
In group 6, in response to PGf2α injection mean P4 concentrations of second sample decreased versus first one (1.07 vs. 2.12 ng mL-1, respectively) then after joint injection of EB and hCG, serum P4 concentration increased in third sample (1.51 ng mL-1). These results show that PGf2α affect primarily on the corpus luteum; then after joint injection of hCG and EB, mean P4 concentration increases. Treatment with hCG is associated with an increase in the number of large luteal cells and a concomitant reduction in the number of small luteal cells in the CL. Administration of hCG induces a greater increase in plasma progesterone concentration compared to GnRH treatment. This could be due to a combined effect on the original and on the induced CL (Xu et al., 1995). In this group, in response to injection of combination of hCG and EB 36 h later (before AI) mean progesterone concentration increased and this demonstrates the effect of hCG and EB on functional CL formation.
Using of two injections of PGf2α with 12 days apart has no pregnancy in the control group in 6 weeks postpartume period, but application of ovsynch in group 2 induced 45 and 50% in estrus and pregnancy rate, respectively; Pursely et al. (1997a) reported use of fixed time AI can result in higher pregnancy rate with no estrus detection. In consistent with Ryan et al. (1999), the results of this experiment show that oestrous detection rate and the synchrony of oestrus was increased after administering oestradiol benzoate in group 3, 5 and 6. Previously, Ryan et al. (1995) reported administration of oestradiol benzoate at after treatment with PGf2α reduced the interval to oestrus and ovulation that can improve pregnancy rate. Particularly we have carried out a short length program of synchronization that show good results than control group. However it seems these findings warrant further investigation.
REFERENCES
- Ahuja, C., F. Montiel, R. Canseco, E. Silva and G. Mapes, 2005. Pregnancy rate following GnRH+PGF2α treatment of low body condition, anestrous Bos taurus by Bos indicus crossbred cows during the summer months in a tropical environment. Anim. Reprod., Sci., 87: 203-213.
CrossRefDirect Link - Bartolome, J., L. Archbald, P. Morresey, J. Hernandez and T. Tran et al., 2000. Comparison of synchronization of ovulation and induction of estrus as therapeutic strategies for bovine ovarian cysts in the dairy cow. Theriogenolog, 53: 815-825.
Direct Link - Britt, J.S. and J. Gaska, 1998. Comparison of two estrus synchronization programs in a large, confinement-housed dairy herd. J. Am. Vet. Med. Assoc., 212: 210-212.
PubMed - Burke, J.M., R.L. De la Sota, C.A. Risco, C.R. Staples, E.J.P. Schmitt and W.W. Thatcher, 1996. Evaluation of timed insemination using a gonadotropin-releasing hormone agonist in lactating dairy cows. J. Dairy Sci., 79: 1385-1393.
Direct Link - Calder, M.D., B.E. Salfen, B. Bao, R.S. Youngquist and H.A. Garverick, 1999. Administration of progesterone to cows with ovarian follicular cysts results in a reduction in mean LH and LH pulse frequency and initiates ovulatory follicular growth. J. Anim. Sci., 77: 3037-3042.
Direct Link - Cavalieri, J., A.R. Rabiee, G. Hepworth and K.L. Macmillan, 2005. Effect of artificial insemination on submission rates of lactating dairy cows synchronised and resynchronised with intravaginal progesterone releasing devices and oestradiol benzoate. Anim. Reprod. Sci., 90: 39-55.
Direct Link - De la Sota, R.L., J.M. Burke, C.A. Risco, F. Moreira, M.A. DeLorenzo and W.W. Thatcher, 1998. Evaluation of timed insemination during summer heat stress in lactating dairy cattle. Theriogenology, 49: 761-770.
CrossRefDirect Link - Fricke, P.M. and M.C. Wiltbank, 1999. Effect of milk production on the incidence of double ovulation in dairy cows. Theriogenology, 52: 1133-1143.
Direct Link - Ferguson, J.D. and D.T. Galligan, 1993. Prostaglandin synchronization programs in dairy herds (Part I). Comp. Contin. Educ. Pract. Vet., 15: 646-655.
Direct Link - Folman, Y., M. Kaim, Z. Herz and M. Rosenberg, 1990. Comparison of methods for the synchronization of estrous cycles in dairy cows. 2. Effects of progesterone and parity on conception. J. Dairy Sci., 73: 2817-2825.
Direct Link - Garverick, H.A., 1997. Ovarian follicular cysts in dairy cows. J. Dairy Sci., 80: 995-1004.
Direct Link - Hobson, W.C. and W. Hansel, 1972. Plasma LH levels after ovariectomy, corpus luteum removal and etradiol administration in cattle. Endocrinology, 91: 185-190.
Direct Link - Khanum, S.A., M. Hussain and R. Kausar, 2006. Manipulation of estrous cycle in Dwarf goat (Capra hircus) using estrumate under different management conditions. Anim. Reprod. Sci., 92: 97-106.
CrossRefDirect Link - Kim, I.H., G.H. Suh and D.S. Son, 2003. A progesterone-based timed AI protocol more effectively prevents premature estrus and incomplete luteal regression than an Ovsynch protocol in lactating Holstein cows. Theriogenology, 60: 809-817.
CrossRefDirect Link - Kristula, M., R. Bartholomew, D. Galligan and C. Uhlinger, 1992. Effects of a prostaglandin F2a synchronization program in lactating dairy cattle. J. Dairy Sci., 75: 2713-2718.
Direct Link - Lopez-Gatius, F., 2000. Reproductive performance of lactating dairy cows treated with cloprostenol, hCG and estradiol benzoate synchronization of estrous followed by timed artificial insemination. Theriogenology, 54: 551-558.
Direct Link - Macmillan, K.L. and A.J. Peterson, 1993. A new intravaginal progesterone releasing device for cattle (DIDR-B) for estrous synchronization, increasing pregnancy rates and the treatment of postpartum anestrous. Anim. Reprod. Sci., 33: 1-25.
Direct Link - Martinez, M.F., J.P. Kastelic, G.P. Adams, B. Cook, W.O. Olson and R.J. Mapletoft, 2002. The use of progestins in regimens for fixed-time AI in beef cattle. Theriogenology, 57: 1049-1059.
Direct Link - Mialot, J.P., F. Constant, P. Dezaux, B. Grimard, F. Deletang and A.A. Ponter, 2003. Estrus synchronization in beef cows: Comparison between GnRH+PGF2+GnRH and PRID+PGF2+eCG. Theriogenology, 60: 319-330.
Direct Link - Michael, J., R. James, E. Kinder, K. Fike, F. Kojima, D. Colin, R. Ogle and C.R. Bunt, 2000. Recent advances in bovine reproductive endocrinology and physiology and their impact on drug delivery system design for the control of the estrous cycle in cattle. Adv. Drug Deliv. Rev., 50: 277-320.
Direct Link - Momcilovic, D., L.F. Archibald, A. Walters, T. Tran, D. Kelbert, C. Risco and W.W. Thatcher, 1998. Reproductive performance of lactating dairy cows treated with gonadotropin-releasing hormone (GnRH) and/or prostaglandin F2α (PGf2α) for synchronization of estrus and ovulation. Theriogenology, 50: 1131-1139.
- Padula, A.M. and K.L. MacMillan, 2002. Reproductive responses of early postpartum dairy cattle to continuous treatment with a GnRH agonist (deslorelin) for 28 days to delay the resumption of ovulation. Anim. Reprod. Sci., 70: 23-36.
Direct Link - Peters, A.R., 2005. Veterinary clinical application of GnRH-questions of efficacy. Anim. Reprod. Sci., 88: 155-167.
PubMedDirect Link - Pursley, J.R., M.O. Mee and M.C. Wiltbank, 1995. Synchronization of ovulation in dairy cows using PGF2α and GnRH. Theriogenology, 44: 915-923.
CrossRefDirect Link - Pursley, J.R., M.R. Kosorok and M.C. Wiltbank, 1997. Reproductive management of lactating dairy cows using synchronization of ovulation. J. Dairy Sci., 80: 301-306.
Direct Link - Pursley, J.R., M.C. Wiltbank, J.S. Stevenson, J.S. Ottobre, H.A. Garverick and L.L. Anderson, 1997. Pregnancy rates per artificial insemination for cows and heifers inseminated at a synchronized ovulation or synchronized estrus. J. Dairy Sci., 80: 295-300.
CrossRefDirect Link - Ryan, D.P., J.A. Galvin and K.J. O'Farrell, 1999. Comparison of oestrous synchronization regimens for lactating dairy cows. Anim. Reprod. Sci., 56: 153-168.
Direct Link - Schmitt, E.J.P., M. Drost, T. Diaz, C. Roomes and W.W. Thatcher, 1996. Effect of a Gonadotropin-Releasing Hormone agonist on follicle recruitment and pregnancy rate in cattle. J. Anim. Sci., 74: 154-160.
Direct Link - Steveson, J.J., Y. Konayashi, M.P. Shipka and K.C. Rauchhotz, 1996. Altering conception of dairy cattle by gonadotropin releasing hormone preceding luteolysis induced by prostaglandin F2α. J. Dairy Sci., 79: 402-410.
Direct Link - Stevenson, J.S., Y. Kobayashi and K.E. Thompson, 1999. Reproductive performance of dairy cows in various programmed breeding systems including Ovsynch and combinations of gonadotropin releasing hormone and prostaglandin F2α. J. Dairy Sci., 82: 506-515.
Direct Link - Tebble, J.E., M.J. O`Donnell and H. Dobson, 2001. Ultrasound diagnosis and treatment outcome of cystic ovaries in cattle. Vet. Rec., 148: 411-413.
Direct Link - Thatcher, W.W., M. Drost, J.D. Savio, K.L. Macmillan and K.W. Entwistle et al., 1993. New clinical uses of GnRH and its analogues in cattle. Anim. Reprod. Sci., 33: 27-49.
Direct Link - Thatcher, W.W., F. Moreina, S.M. Pancarci, J.A. Bartolome and J.E.P. Santos, 2002. Strategies to optimize reproductive efficiency by regulation of ovarian function. Domest. Anim. Endocrinol., 23: 243-254.
Direct Link - Todoroki, J., H. Yamakuchi, K. Mizoshita, N. Kubota and N. Tabara et al., 2001. Restoring ovulation in beef donor cows with ovarian cysts by progesterone-releasing intravaginal silastic devices. Theriogenology, 55: 1919-1932.
Direct Link - Trigg, T., 2005. Applications of GnRH in the control and management of fertility in female animals. Anim. Reprod. Sci., 88: 141-153.
Direct Link - Twagiramungu, H., L.A. Guilbault and J.J. Dufour, 1995. Synchronization of ovarian follicular waves with a gonadotropin-releasing hormone agonist to increase the precision of estrus in cattle: A review. J. Anim. Sci., 73: 3141-3151.
Direct Link - Webb, R., B.K. Campbell, H.A. Garverick, J.G. Gong, C.G. Gutierrez, and D.G. Armstrong, 1999. Molecular mechanisms regulating follicular recruitment and selection. J. Reprod. Fertil. Suppl., 54: 33-48.
Direct Link - Xu, Z.Z., L.J. Burton and K.L. Macmillan, 1995. Reproductive performance of lactating dairy cows following estrous synchronization regimens with Pgf2α and progesterone. Theriogenology, 47: 687-701.
Direct Link