
S. H. Ghorbani
Department of Animal Breeding and Genetics,
Animal Science Research Institution of Iran (ASRI), Karaj, Islamic Republic of Iran
M. A. Kamali
Department of Animal Breeding and Genetics,
Animal Science Research Institution of Iran (ASRI), Karaj, Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3215-3219







ABSTRACT
Genetic parameters were estimated in base population of a closed experimental strain fowl, from data issued from 13 successive generations of selection. This population had been selected for body weight at 12 weeks of age (BW12) and egg number during the first 12 weeks of laying period (EN), mean egg weight at 28th, 30th, 32nd weeks and Age at Sexual Maturity (ASM). Data were obtained on 35461 Iranian native hens belonging to breeding center for Fars province in Iran. The method of multi-traits restricted maximum likelihood with an animal model was used to estimate genetic parameters. Resulting heritabilities for BW12, EN, EW and ASM were 0.58, 0.34, 0.62 and 0.49, respectively. Genetic correlations between BW12 and EN, EW and ASM were -0.06, 0.49 and 0.02, respectively. Genetic correlations between EN and EW and ASM were -0.26 and-0.77, respectively, while between EW and ASM, it was 0.20. The overall predicted genetic gains, after 13 generations of selection, estimated by the regression coefficients of the breeding value on generation number were equal to 9.55, 0.99, 0.05 and -1.66, for BW12, EN, EW and ASM, respectively.
PDF Abstract XML References Citation
How to cite this article
S. H. Ghorbani and M. A. Kamali, 2007. Genetic Trend in Economic Traits in Iranian Native Fowl. Pakistan Journal of Biological Sciences, 10: 3215-3219.
DOI: 10.3923/pjbs.2007.3215.3219
URL: https://scialert.net/abstract/?doi=pjbs.2007.3215.3219
DOI: 10.3923/pjbs.2007.3215.3219
URL: https://scialert.net/abstract/?doi=pjbs.2007.3215.3219
INTRODUCTION
Iranian native hens are valuable genetic resources, due to their adaptability to harsh conditions in rural area. The experimental flock from Fars province gene pool has been established in 1987. Relationships among growth and other economic traits for chickens have been reviewed under short-and long-term selection for chickens (Liu et al., 1994, 1995; Argentao et al., 2002; Rance et al., 2002; Gaya et al., 2006), turkeys (Buss, 1990; Nestor et al., 2000), ducks (Hu et al., 1999, 2004; Kleczek et al., 2006), native fowls in Iran (Kamali et al., 1995; Esmaeilkhanian et al., 2004). Kamali et al. (1995) has been used, different methods of selection index for selection of economic traits in Iranian indigenous hens. On the other hand, most of the estimates have been obtained through traditional estimation methods, but only very few of these were based on Restricted Maximum Likelihood (REML) methodology (Patterson and Thompson, 1971) and animal models (Henderson, 1984). Best Linear Unbiased Prediction (BLUP) in animal models (Henderson, 1988) has been used more and more for predicting the genetic merit of candidates for selection in most species of farm animals. For this purpose estimates of genetic parameters in the base population are required. Also, the effects of selection for growth rate on laying traits were not known in Iranian native hens. Therefore, it was useful to evaluate whether selection for growth rate in juvenile fowls impairs laying traits and whether selection for egg production is possible or not. Thus, the goal of this study was to estimate the heritabilities, genetic correlations and breeding values of 4 traits (body weight at 12 weeks of age, egg number during first 12 weeks of laying period, mean egg weight at 28th, 30th and 32nd weeks and age at sexual maturity) in native fowls of Fars province in Iran. The estimated genetic parameters were used to calculate predicted genetic trends over 13 generations of selection. Because, it was necessary, to evaluate whether these economic traits, over 13 generation of selection with multivariate REML method, improved or not.
MATERIALS AND METHODS
Birds and data: Fars province in Iran is situated between latitudes 27°3/ to 31°40/ N and longitude 50°36/ to 55°35/ E and benefits from a moderate climate with averaged monthly temperatures of 6 to 13°C in winter and 27 to 29°C in summer.
In this experiment, data on 35461 Iranian native hens belonging to Breeding Center for Fars Native Chicken, from first Generation (G1), to Generation 13 (G13) were used to estimate the genetic parameters of growth and production traits. The performance records of female and male birds, collected between 1988 to 2005. All laying native fowls were from a small population selected for individual phenotypic value of body weight at 12 weeks of age (BW12) and Egg Numbers (EN) during the first 12 weeks of laying period. In the first generation, eggs were randomly collected from rural areas without any history of introduction of exotic birds and were hatched to constitute the base population. Parents were selected on the basis of BW12 and EN. Cocks were selected on the basis of BW12 and the production of their sisters. Day-old chicks were wing-banded. All birds were reared on floor, intermingled during the rearing period for 12 weeks. After 12 weeks of age, the laying native fowls, selected according to body weight at 12 weeks of age in the first selection step, were then transferred into individual cages and the production traits were recorded for 12 weeks. Average selection intensities of about 40% for hens and 5% for cocks were realized during the experiment.
Animals were fed with a diet containing 175 g CP kg-1 and 2900 kcal kg-1 ME during the growing period and a diet containing 160 g CP kg-1 and 2900 kcal kg-1 ME during the laying period. Drinking water was provided ad libitum throughout the experiment. Individual data were calculated for 4 laying traits: Body weight at 12 weeks of age (BW12), Egg Number (EN) during the first 12 weeks of laying period, mean egg weight at 28th, 30th and 32nd weeks (EW) and Age at Sexual Maturity (ASM).
Statistical analyses: Estimates of heritability, genetic correlations and breeding values were based on the animal model procedure. Variance-covariance components were estimated using a multivariate animal model with Derivative-Free Restricted Maximum Likelihood (DF REML) method (Meyer, 1997). Fixed effects were generation, hatch and sex and random effects were animal and residual.
The following model of analysis was used:
yi = Xi bi + Zi ai + ei |
where yi is the vector of observations of ith trait; X and Z are incidence matrices relating observations to the respective fixed and random effect; bi is a vector of fixed effects on observations of ith trait; ai is vector of random additive genetic effect for ith trait; ei is a vector of random residual effects for ith trait and I = 1,…., 4 consists of BW12, EN, EW and ASM. The vector b1 consists of fixed effects of the combined Generation and Hatch (GH) effect and of sex effect on body weight. The vector b2 consists of fixed effects of the combined Generation and Hatch (GH) effect and of the number of recording days on egg number (covariate). Vectors b3 and b4 consist of fixed effect of the combined Generation and Hatch (GH) effect that affected average egg weight and age at sexual maturity. The vectors a1 to a4 consist of random effect of breeding values of above traits, respectively. In this model, the environmental and genetic maternal effects are incorporated into the residual.
The general linear model procedure was used to calculate the mean, standard error and least square analysis of variance for each trait. The phenotypic and genetic trends for BW12, ASM, EN and EW were estimated by regression of average Least Square Means (LSM) and regression of average breeding values on the generation number, respectively. The graphs for average phenotypic trend of each trait across the generations were obtained by drawing the least square means across generations and average genetic trend of each trait across the generations was obtained by drawing the average breeding values across generations.
RESULTS
Results obtained from analysis of variance of traits (GLM) showed that hatchability and generation had significant effect on traits (p<0.0001). Sex and the number of recording days of egg number (covariate) were found to have a significant effect on BW12 and EN, respectively. Figure 1a-d reports phenotypic trends and Fig. 2a-d genetic trends for BW12, ASM, EN and EW, respectively. The regression coefficient of least square means on generation number were equal to 17.94, -2.81, -1.10 and 0.14, for BW12, ASM, EN and EW, respectively. They were significant for all of these traits except for EN. For this reason there are reasonable change in production of traits compare to generation one except for EN (Fig. 1) The total means of BW12, ASM, EN and EW were 835.87 g, 164.85 day, 52.36 eggs and 43.96 g, respectively.
As shown in Table 1, estimated heritabilities for BW12, EN, EW and ASM, were equal to 0.58±0.02, 0.34±0.02, 0.62±0.02 and 0.49±0.02, respectively. Phenotypic correlations between BW12 and EN and EW were relatively low (0.05 and 0.34, respectively) but it was negative with ASM (-0.10). The genetic correlations between BW12 and EN and ASM were also very low (-0.06, 0.02, respectively), but it was relatively high and positive with EW (0.49).
| Table 1: | Heritabilities, genetic and phenotypic correlations among the 4 economic traits |
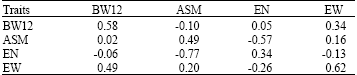 | |
| BW12 = Body weight at 12 weeks of age, ASM = Age at sexual maturity, EN = Egg number during the first 12 weeks of laying period, EW = Mean egg weight at 28th, 30th, 32nd weeks, Heritabilities are given on the diagonal, genetic correlations below diagonal, Phenotypic correlations above diagonal | |
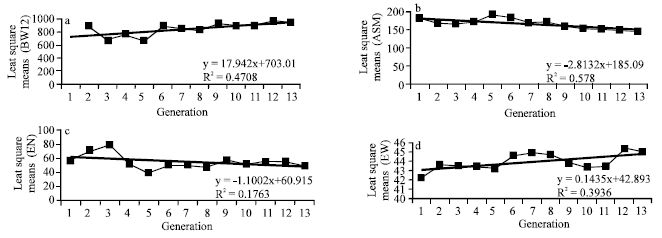 | |
| Fig. 1: | Phenotypic trend of: (a) body weight at twelve week age (BW12), (b) Age at sexual maturity (ASM), (c) Egg number (EN) and (d) Average egg weight (EW) in 13 generation selection |
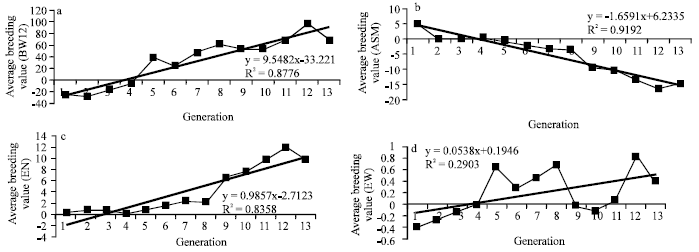 | |
| Fig. 2: | Genetic trend of: (a) body weight at twelve week age (BW12), (b) Age at sexual maturity (ASM), (c) Egg number (EN) and (d) Average egg weight (EW) in 13 generation selection |
Genetic and Phenotypic correlations between ASM and EN were high and negative (-0.77 and-0.57, respectively), while they were low between ASM and EW (0.20 and 0.16, respectively). Genetic and Phenotypic correlations between EN and EW were low (-0.26 and-0.13, respectively).
The regression coefficient of average breeding value for BW12 on generation number was positive (9.55). The cumulated genetic change for BW12 increased from G1 to G13. Regression coefficient of average breeding values for ASM on generation number was negative (-1.66). Egg production exhibited relatively irregular trend between G1 to G13. The regression coefficient was positive and low (0.99) and significant. Egg weight exhibited irregular trend between G1 to G13, except in the first four generation, where it had positive trend that confirmed progress in this trait. The regression coefficient was positive and very low (0.05) but significant.
DISCUSSION
The results of the present study showed that selection had modified growth and laying traits. The models proposed by Meyer (1992) to estimate the variances and covariances for the common environmental maternal effects or for the maternal genetic effects were not used. Considering the total amount of available information and the data structure, the parameters would not have been correctly estimated in a complete model due to over-parameterization (Chapuis et al., 1996). The relative influence of maternal effects is generally moderate in poultry. In chickens, this proportion varied from 3 to 8% depending on the trait, for body weight, breast muscle weight and abdominal fat weight (Le Bihan-Duval et al., 1998). It was preferred to use a simple model with additive direct genetic effects, neglecting the maternal effects. The maternal effects in poultry can be explained by the expected influences of egg characteristics on the development of the embryo and the early growth of the bird. When neglecting the environmental maternal effects, Le Bihan-Duval et al. (1998) found in chickens that the direct heritability was overestimated, on average over both sexes by 12 to 19% according to the trait. The present analysis could have overestimated the heritability estimates. The relatively high value of heritability estimated for these traits could be also due to shorter selection periods in this flock. The main reason for high absolute values of heritability in this flock can be attributed to the large size of base population with wide dispersal environments and low amount of inbreeding (Falconer, 1996). Kamali et al. (1995) obtained in the same flock for one generation and by using nested design a lower heritability for BW12 and EW but higher for ASM. Nevertheless the simple model with animal genetic effect as the only random effect corresponds to the model likely to be used in routine evaluation.
The negative genetic correlation between Egg Number (EN) and body weight at 12 weeks of age (BW12) shows that improvement in one trait is not detrimental to the other trait. These results were same from those reported by Hagger (1994) for crosses between Rhode Island Red cocks and Leghorn hens. In their study, the net effects of genetic increases in growth rate of juvenile body weight on female reproduction rate were negative. A positive relationship was apparent between body weight and ova production. In long term selection experiments, onset of maturity in low weight line pullets may even be inhibited. These chickens fail to attain the necessary body weight or composition (Crawford, 1993).
We do not find significant genetic correlation between EN and EW, but Fairfull and Gowe (1990) in unselected control lines of White Leghorns reported the positive correlations and Besbes et al. (1992), Mielenz et al. (1994), Wei and Van der Werf (1993), Hagger (1994), Tixier-Boichard et al. (1995), Jeyarubau and Gibson (1996) and Richard et al. (2006) reported negative correlations. High levels of heritability of BW12 and EW as well as their positive genetic correlation showed that selection for increasing body weight induced increasing in average Egg Weight (EW). Egg production is genetically and Phenotypic negatively correlated with age at sexual maturity (-0.77; -0.57, respectively). The study of Poggenpoel et al. (1996) reported similar result for genetic correlation between these traits, result was reported by Liu et al. (1995) for low-weight selected line is different from those that. In their experiment negative correlation found for these traits in high-weight selected line. Present study highlighted a high negative environmental and genetic correlation between ASM and EN, both traits having significant levels of heritability. This shows that selection for increased egg production may decrease the ASM and in long term selection program lead to detrimental effect from a physiological point of view. In this study, genetic progress was achieved. The low genetic correlation between BW12 and EN indicated opportunity for future improvement of both traits in such a population. Selection for BW12 and EN should not impair improvement of ASM and EW. More research is needed to improve the selection method of mating system to avoid inbreeding depression.
ACKNOWLEDGMENT
This research was carried out at Department of Animal Breeding and Genetic, Animal Science Research Institution of Iran and Department of livestock affairs, Ministry of Jihad-e-Agriculture.
REFERENCES
- Argentao, C., T. Michelan Filho, J.B. Marques, E.M. Souza, J.P. Eler and J.B.S. Ferraz, 2002. Genetic and phenotypic parameters of growth and carcass traits of a male line broilers raised in tropical conditions. Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, August 19-23, 2002, Montpellier, France, pp: 333-336.
- Besbes, B., V. Ducrocq, J.L. Foulley, M. Protais, M. Tavernier, M.T. Boichard and C. Beaumont, 1992. Estimation of genetic parameters of egg production traits of laying hens by restricted maximum likelihood applied to a multiple-trait reduced animal model. Gen. Sel. Evol., 24: 539-552.
CrossRef - Chapuis, H., M. Tixier-Boichard, Y. Delabrosse and V. Ducrocq, 1996. Multivariate restricted maximum likelihood estimation of genetic parameters for production traits in three selected turkey strains. Gen. Selection Evol., 28: 299-317.
CrossRef - Esmaeilkhanian, S., R. Osfori, M.A. KamalI and P. Horn, 2004. Genetic variation among native chicken breeds of Iran based on biochemical polymorphisms. Br. Poul. Sci., 45: 598-603.
Direct Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Gaya, L.G., J.B.S. Ferraz, F.M. Rezende, G.B. Mourao, E.C. Mattos, J.P. Eler and T.F. Michelan, 2006. Heritabilities and genetic correlation estimates for performance and carcass and body composition traits in a male broiler line. Poul. Sci., 85: 837-843.
Direct Link - Hu, Y.H., J.P. Poivey, R. Rouvier, S.C. Liu and C. Tai, 2004. Heritabilities and genetic correlation of laying performance in-Muscovy ducks selected in Taiwan. Br. Poul. Sci., 45: 180-185.
Direct Link - Jeyarubau, M.G. and J.P. Gibson, 1996. Estimation of additive genetic variance in commercial layer poultry and simulated populations under selection. Theor. Applied Gen., 92: 483-491.
CrossRefDirect Link - Kamali, M.A., S. Toth and I. Szalai, 1995. Development of selection indices for indigenous hens of Iran. Allattenyesztes es Takarmanyozas, 44: 495-506.
Direct Link - Kleczek, K., K. Wawro, O.E. Wilkiewicz-Wawr and W. Makowski, 2006. Multiple regression equations to estimate the content of breast muscles, meat and fat in Muscovy ducks. Poul. Sci., 85: 1318-1326.
CrossRefPubMedDirect Link - Le Bihan-Duval, E., S. Mignon-Grasteau, N. Millet and C. Beau-Mont, 1998. Genetic analysis of a selection experiment on increased body weight and breast muscle weight as well as on limited abdominal fat weight. Br. Poult. Sci., 39: 346-353.
PubMed - Liu, G., E.A. Dunnington and P.B. Siegel, 1995. Correlated responses to long term divergent selection for eight-week body weight in chickens: Growth, sexual maturity and egg production. Poul. Sci., 74: 1259-1268.
PubMedDirect Link - Nestor, K.E., J.W. Erson and R.A. Patterson, 2000. Genetics of growth and reproduction in the turkey. 14. Changes in genetic parameters over thirty generations of selection for increased body weight. Poul. Sci., 78: 445-452.
CrossRefDirect Link - Nestor, K.E., J.W. Erson, R.A. Patterson and S.G. Velleman, 2006. Genetics of growth and reproduction in the Turkey. 16. Effect of repeated backcrossing of an egg line to a commercial sire line. Poul. Sci., 85: 1550-1554.
Direct Link - Poggenpoel, D.G., G.F. Ferreira, J.P. Hayes and J.J. Du Preez, 1996. Response to long-term selection for egg production in laying hens. Br. Poul. Sci., 37: 743-756.
CrossRefDirect Link - Rance, K.A., G.M. Mcentee and R.M. Mcdevitt, 2002. Genetic and phenotypic relationships between and within support and demand tissues in a single line of broiler chicken. Br. Poul. Sci., 43: 518-527.
PubMedDirect Link - Richard, M.M., C. Marie-Etancelin, F. Dubos, M.D. Bernadet, G. Guy and J.M. Brun, 2006. Genetic parameters of reproduction and force-feeding traits in geese under artificial lighting conditions. Proceedings of the 8th Congeress on Genet. Applied Livest. Prod., August 13-18, 2006, Belo Horizonte, Brasil, pp: 213-218.