
L. Fekrat
Department of Plant Protection, Faculty of Agriculture, Shahid Chamran University, Ahwaz, Iran
P. Shishehbor
Department of Plant Protection, Faculty of Agriculture, Shahid Chamran University, Ahwaz, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3180-3184







ABSTRACT
Development and reproduction of the cotton whitefly, Bemisia tabaci (Gennadius) were studied on aubergine, tomato and potato under laboratory conditions (30°C and 55% RH). Total life cycle from egg to adult was 14.9, 20.0 and 14.2 days on aubergine, tomato and potato, respectively. Immature mortality were 12.9, 18.1 and 12.3% at the same three host plants. Females of B. tabaci oviposited means of 51.8, 60.1 and 67.5 eggs on aubergine, tomato and potato, respectively and had a mean longevity of 8, 14 and 12.9 days on the same three host plants. The net reproductive rate was 18.12, 15.06 and 27.63 and the daily intrinsic rate of increase was 0.141, 0.092 and 0.165 on aubergine, tomato and potato, respectively.
PDF Abstract XML References Citation
How to cite this article
L. Fekrat and P. Shishehbor, 2007. Some Biological Features of Cotton Whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) on Various Host Plants. Pakistan Journal of Biological Sciences, 10: 3180-3184.
DOI: 10.3923/pjbs.2007.3180.3184
URL: https://scialert.net/abstract/?doi=pjbs.2007.3180.3184
DOI: 10.3923/pjbs.2007.3180.3184
URL: https://scialert.net/abstract/?doi=pjbs.2007.3180.3184
INTRODUCTION
The cotton whitefly, Bemisia tabaci (Gennadius), is one of the most important agricultural insect pests in the middle east, Europe, North and Central America (Mound and Halsey, 1978; Lopez-Avilla, 1986; McKenzie et al., 2004). In addition to feeding on more than 700 host plant species within 86 botanical families (Greathead, 1986), B. tabaci has a high reproductive capacity and destructive life habits that enable it to (1) cause severe damage through plant feeding and (2) transmit more than 90 types of virus diseases in commercial crops (Brunt, 1986; Morales and Jones, 2004).
Several studies on the biology of B. tabaci have been conducted under diverse environmental conditions (Lopez-Avilla, 1986; Coudriet et al., 1985; Hendi et al., 1984; Powell and Bellows, 1992a, b; Xu et al., 2003; Al-Zeyoud and Sengonca, 2004). Those studies reported that the life cycles varied mainly depending upon the temperature, relative humidity and the host plant. Russell (1975) compiled literature on the biology and morphology of B. tabaci and other whitefly species in legume crops. She reported finding much variability in the life cycle and other biological aspects that were strongly related to climatic factors and the host plant.
Host plant races are one variant of biotype (Diel and Bush, 1984) of B. tabaci, now referred to as biological types, that exhibit variation in geographical distribution, host range, fecundity, dispersal behavior, insecticide resistance, natural enemy complexes and endosymbiont complement (Rowland et al., 1991; Costa et al., 1993a, b, 1995; Bedford et al., 1994; Kirk et al., 2000). Therefore, for each pest management program an exact determination of the crucial population parameters is required. This study was conducted to determine the development and reproduction of B. tabaci under controlled climatic conditions on different solonaceous vegetable crops.
MATERIALS AND METHODS
Stock culture maintenance: Adult cotton whitefly, Bemisia tabaci used in these studies were collected in September 2004 from an aubergine field in Shahid Chamran University Campus, Ahwaz, Iran and reared on the foliage of aubergine plants (cultivar Blackbeauty) grown from seeds and transplanted into compost in plastic pots (20 cm diameter). Infested plants were kept in wooden-framed rearing cages (120x60x60 cm) covered with white nylon mesh of 210 μm aperture. They were maintained in a laboratory where seasonal temperature ranged from 16-25°C and relative humidity 40-50%. The photoperiod was 12 : 12 (L : D) h, with illumination (4000 lux) provided from fluorscent lamps. Plants were kept in the cages until they were severly damaged by the whiteflies; new plants being added when needed.
Three host plant species were used in the study are tomato, Lycopersicon esculentum, cultivar Octaplus; aubergine, Solanum melongena, cultivar Blackbeauty; potato, Solanum tubersum, cultivar Cosima.
Developmental time and mortality: The effect of different host plant species on developmental time and mortality of B. tabaci was studied by confining 30-50 adult whitefiles of different ages on the undersurface of different plant leaves by means of a clip cage as described by Costa et al. (1991). After 24 h, the whiteflies and cages were removed and the test plants harbouring eggs were immediately transferred to a growth chamber (30±1°C). Relative humidity ranged from 50 to 60 %, a photoperiod of 14 : 10 (L : D) h was maintained and the light intensity was 4000 lux. The development of immature stages was monitored with a dissecting microscope until adult emergence. The duration of immature stages and mortality were recorded daily. The percentage mortality of each immature stage was calculated as:
Longerity, fecundity, sex ratio and life tabe parameters: The effect of different host plants on longevity, fecundity, sex ratio and other population parameters was studied by confining one adult female and one male immediately on emergence on the undersurface of different plant leaves by means of a clip cages similar to those described above. Every other day, whiteflies were transferred to a new leaf and the eggs laid were counted. Each experiment was terminated with the natural death of the female. All eggs were monitored daily until adult eclosion and the numbers of male and female mites were recorded to determine the sex ratio.
Longevity and daily fecundity were used in conjunction with the data on the developmental period and survival of immature stages to construct age specific survival and fecundity tables and to calculate the following: Gross Reproductive Rate (GRR), net reproductive rate (Ro), intrinsic rate of increase (rm), finite rate of increase (λ), mean generation time (T) and population Doubling Time (DT) (Birch, 1948; Andrewartha and Birch, 1954; Southwood, 1978; Krebs, 1985). The sex ratio of 1 female : 1 male was used to calculate life table parameters.
Analysis of variance (ANOVA) and Duncan multiple range test were used to determine the effect of host plant on developmental duration, longevity, fecundity and sex ratio of whiteflies. A series of Chi-square tests were conducted to determine if there were any significant differences in stage mortality for insects reared on different host plants.
RESULTS AND DISCUSSION
Development time: Results in this study showed that the time required for B. tabaci to complete development from egg to adult was influence by the host plant on which the whitefly was reared. The time required for B. tabaci to complete development in the laboratory from egg deposition to adult eclosion on three different crops is shown in Table 1. The developmental duration times from egg to adult were 14.9, 20.0 and 14.2 days on aubergine, tomato and potato, respectively. The minimum developmental time was 13.5 days on potato and the maximum time was 21.5 on tomato. About 25% less time was required for B. tabaci to complete life cycle on aubergine and potato than on tomato. Analysis of variance indicated significant differences in development time between the host plants examined (p = 0.0000).
These data are in line with the results of Hendi et al. (1984) who reported 19.5 days for developmental duration of B. tabaci on tomato at 30°C. Butler et al. (1983) observed developmental time of 17 days on cotton at 30°C which were higher than those recorded in this study on aubergine (14.9 day) and potato (14.2 day). Powell and Bellows (1992a) observed developmental times of 17.64 and 16.74 days on cotton and cucumber, at 30°C (through interpolation) for B. tabaci which were longer than those recorded in this study on aubergine and potato. The difference may be explained by disparities in host plant suitability or population differences between the whiteflies.
Mortality: Mean total mortality ranged from 12-18%. Mortality in the various developmental stages varied from 0.0 to 7.5% (Table 2). Stage mortality examined between host plant species (p<0.05; χ2 = 5.99).
| Table 1: | Developmental time in days (Mean±SD) of B. tabaci on different host plants |
 | |
| Means in the each row followed by the same letter were not significantly different at the 0.05 level when tested by Duncan multiple range test; Sample size (n) in parentheses | |
| Table 2: | Percentage mortality within stage of B. tabaci on different host plants |
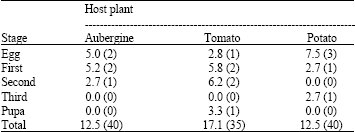 | |
| Sample size (n) in parentheses is No. dying in each stage except for total which is the initial No. entering the egg stage | |
There were no significant mortality differences between host plants for egg, nymphal and pupal stages (egg ( χ2 = 0.822), first nymphal stage (χ2 = 0.473), second nymphal stage (χ2 = 2.366), third nymphal stage (χ2 = 1.855) and pupal stage (χ2 = 2.356)).
Total mortalities in the current study differ from those reported by other researchers in the previous studies. Total mortality of B. tabaci was reported to be 50% (on tomato at 30°C (Hendi et al., 1984), 45 and 35% on cotton and cucumber, respectively, at 29°C; Powell and Bellows, 1992a).
Longevity, fecundity, sex ratio and life table parameters: Longevity varied with host plants. Mean longevity of females ranged from 8 days on aubergine to 14 days on tomato (Table 3). An ANOVA showed that the longevities of females on different host plants were significantly different (p = 0.0048). The longest lived individuals survived for 22 days on tomato and the shortest were whiteflies that survived for only 6 days on aubergine. The longevities of male were significantly different overall (p = 0.009).
The longevities of adult B. tabaci determined in the present study were similar to those reported in other studies conducted at similar constant temperatures (Butler et al., 1983; Powell and Bellows, 1992b). Butler reported that at 26.7 and 32°C females lived 8 and 10.4 days and males 7.6 and 11.7 days on cotton. Powell and Bellows (1992c) reported that females lived 9.6 days and males 7.03 days at 29°C on cucumber. As in the present study, most researchers reported that female insects lived longer than males.
Information on fecundity is crucial in determining potential insect populations on different host plants. Cotton whitfly fecundity was greatly affected by host plant. The whiteflies which produced the greatest and least number of eggs, on average, were those reared on potato and aubergine with 67.5 and 51.8 eggs, respectively (Table 3). Whitefly reared on tomato produced the most eggs with 95 being produced in 22 days. The largest mean egg/day value was for whiteflies reared on aubergine with 5.8 eggs/day. The least were 4.2 eggs/day laid on tomato. ANOVA indicated no significant host plant effects on mean total fecundity (p = 0.212) and mean daily fecundity (p = 0.166).
Other laboratory studies have reported a variety of fertility values for this species. At 29°C Powell and Bellows (1992b) found that B. tabaci laid 73.7 and 208.6 eggs on cotton and cucumber, respectively. Hendi et al. (1984) reported on tomato plants at 30°C a mean of 203.1 eggs. Horowitz (1983) reported a mean of 95.5 eggs laid on cotton at 30°C.
| Table 3: | Longevity (in days), Fecundity, sex ratio and life table parameters of B. tabaci on different host plants |
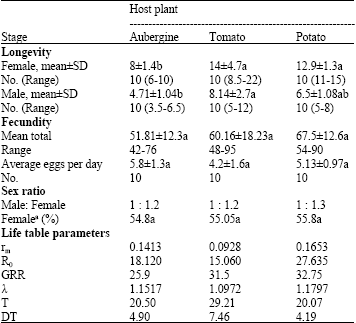 | |
| Means in the each row followed by the same letter were not significantly different at the 0.05 level when tested by Duncan multiple range test; aNo. females/total emerged | |
By comparison in the present study on aubergine, tomato and potato respectively at 30°C, the mean total eggs laid were 51.8, 60.1 and 67.5, generally slightly lower than most other reports.
The mean number of eggs laid per day in the present study may be compared with a study conducted by Hendi et al. (1984) on tomato. He found that the mean number of eggs laid per day was 8, with a range of from 5.5 to 10.8. A partial explanation for the lower number of eggs being laid daily on tomato in the present study, may be due to differences in tomato cultivars used. Hendi et al. (1984) did not state the cultivar that he used. The tomato cultivar used in our experiment (Blackbeauty) may be moderately resistant to B. tabaci. Another explanation may be the population differences in the whiteflies and perhaps to differences in the lighting condition (Hendi did not state the photoperiod under which he conducted his experiment).
Results of this study showed that the sex ratio of cotton whitefly was not influenced by host plant on which the whiteflies were reared. Analysis of variance indicated no significant differences in sex ratio between the host plants examined (p = 0.313) (Table 3).
The species of host plant did not influence the sex ratio of B. tabaci which was female biased on all host plants examined. These data are in agreement with the results of van Lenteren and Noldus (1990) who found that sex ratio of the glasshouse whitefly, Trialeurodes vaporariorum (Westwood) is independent of host plant species.
All life table statistics varied with host plant (Table 3). Fecundity and survivorship data reflect the highest GRR of B. tabaci on potato and the lowest on aubergine. The intrinsic rate of increase (rm) is an instantaneous growth rate of a population under a given condition. Hence, the interpretation of this statistic could be better made by its discrete analogue, the finite rate of increase (λ). The finite rate of increase is the capacity for population growth per unit time. Host plant had a pronounced effect on finite rate of increase (Table 3). However, B. tabaci was found to be capable of population increase (λ> 1) on all host plant tested (Table 3). As shown, the finite rate of increase of this species ranged from λ = 1.17 for those reared on potato to λ = 1.09 for those reared on tomato. The highest rate of population increase occurred on potato and aubergine whereas tomato yielded the lowest rate of population increase. Mean generation time (T) was also influenced by host plant. Tomato and potato had the longest (29.2 days) and shortest (20 days) generation time, respectively. The Doubling Time (DT) value on tomato was about two times that of potato and this suggests that B. tabaci population would triple on potato by the time it would double on tomato.
The results of these investigations indicated that potato and aubergine were the most suitable host plant for development and reproduction of B. tabaci, with tomato least suitable. Hence, one should consider the susceptibility of the host plants when designing pest management program for the B. tabaci in southwestern of Iran.
REFERENCES
- Bedford, I.D., R.W. Briddon, J.K. Brown, R.C. Rosell and P.J. Markham, 1994. Geminivirus transmission and biological characterisation of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Applied Biol., 125: 311-325.
CrossRefDirect Link - Birch, L.C., 1948. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol., 17: 15-26.
CrossRefDirect Link - Costa, H.S., D.M. Westcot, D.E. Ullman, R. Rosell, J.K. Brown and M.W. Johnson, 1995. Morphological variation in Bemisia endosymbionts. Protoplasma, 189: 194-202.
CrossRefDirect Link - Diehl, S.R. and G.L. Bush, 1984. An Evolutionary and applied perspective of insect biotypes. Ann. Rev. Entomol., 29: 471-504.
CrossRef - Kirk, A.A., L.A. Lacey, J.K. Brown, M.A. Ciomperlik, J.A. Goolsby, D.C. Vacek and L.E. Wendel, 2000. Variation within the Bemisia tabaci species complex (Hemiptera Aleyrodidae) and its natural enemies leading to successful biological control of Bemisia biotype B in the USA. Bull. Entomol. Res., 90: 317-327.
Direct Link - McKenzie, C.L., P.K. Anderson and N. Villareal, 2004. An extensive survey of Bemisia tabaci (Homoptera: Aleyrodidae) in agricultural ecosystems in Florida. Florida Entomol., 87: 403-407.
Direct Link - Morales, F.J. and P.J. Jones, 2004. The ecology and epidemiology of whitefly-transmitted viruses in Latin America. Virus Res., 100: 57-65.
CrossRef - Powell, D.A. and T.S. Bellows, Jr., 1992. Adult longevity, fertility and population growth rates for Bemisia tabaci (Genn.) (Hom. Aleyrodidae) on two host plant species. J. Applied Entomol., 113: 68-78.
Direct Link - Rowland, M.B., B. Ackett and M. Stribley, 1991. Evaluation of insecticide in field-control simulators and standard laboratory bioassays against resistant and susceptible Bemisia tabaci (Homoptera: Aleyrodidae) from Sudan. Bull. Entomol. Res., 81: 189-199.
Direct Link