
Fatemeh Abedini
Razi Serum Research Institute, Karaj, Iran
Tahereh Foroutan
Department of Biology, Faculty of Basic Science, Tarbiat Moalem University, Tehran, Iran
Leila Jahangiri
School of Pharmacy, Mashbad University of Medical Sciences, Mashhad, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3146-3149







ABSTRACT
Endothelial progenitor cells from the pulp of milk teeth were isolated for use in clinical applications and tissue engineering. Normal deciduous teeth from children of 7 to 8 years of age, which more than half the tooth root was extracted, were selected from the dental centre. Cells from enzyme treated pulps were cultured and cells resulting from the fifth and eight subculture were combined for cell surface marker determination experiments. Cells were positive for CD34 marker with a total of 99/45%, determined by flowcytometry. Cells also demonstrated alkaline phosphatase (ALP) activity. From the developmental point of view, stem cells from the dental pulp seem to have derived from the neural crest, which our findings technically support this theory. In essence mobile progenitor cells from bone marrow of endothelial origin could also play a significant role in the derivation of dental pulp stem cells.
PDF Abstract XML References Citation
How to cite this article
Fatemeh Abedini, Tahereh Foroutan and Leila Jahangiri, 2007. Alkaline Phosphatase and CD34 Reaction of Deciduous Teeth Pulp Stem Cells. Pakistan Journal of Biological Sciences, 10: 3146-3149.
DOI: 10.3923/pjbs.2007.3146.3149
URL: https://scialert.net/abstract/?doi=pjbs.2007.3146.3149
DOI: 10.3923/pjbs.2007.3146.3149
URL: https://scialert.net/abstract/?doi=pjbs.2007.3146.3149
INTRODUCTION
Postnatal stem cells have been isolated from a variety of tissues including but not limited to bone marrow, brain, skin, hair follicle, skeletal muscle and dental pulp (Spradling and Durmmond Dkai, 2001; Grontho et al., 2000) recently the extraordinary plasticity of postnatal stem cells has been suggested, in which neral stem cells may contribute to blood and skeletal muscle (Gronthos et al., 2000; Miura et al., 2003) bone marrow stem cells may contribute to muscle, liver and neuronal tissue (Parnner et al., 2002; Brazelton et al., 2000). Recent emerging evidence suggested that cell-fusion events may account for some of these observations (Ying et al., 2000). It is necessary to gain further insight into the characteristics of postnatal stem cells and examine their full development potential in vivo (D'Amour and Gage, 2002).
The transition from deciduous teeth to adult permanent teeth is a very unique and dynamic process in which the development and eruption of permanent teeth coordinate with the resorption of the roots of deciduous teeth. Ex vivo expanded stem cells from human exfoliated deciduous teeth were found to express the cell-surface molecules STRO-1 and CD146, 2 early mesenchymal stem cells markers previously found to be present in bonemarrow stem cells (Miura et al., 2003). STRO-1 and CD146 positive cells were found to be located around blood vessele of the remnant pulp, implying that stem cells from human exfoliated deciduous teeth may have originated from a perivasclar microenvironment. In this study we isolated a distinctive population of multipotent stem cells from pulp of exfoliated deciduous teeth and then emphasized also on the markers of endothelial cell line.
MATERIALS AND METHODS
This study was done in Razi serum research institute, Karaj, Iran and Center of research of cellular and molecular-faculty of medical of Beheshti University, Tehran, Iran.
Cells and culture: Deciduous teeth with half extracted roots were obtained from dental centre and were immediately washed in PBS. The dental, pulp was separated by an excavator and incubated in a trypsin solution in 4°C overnight. Isolated cells were passed to a 70 μm filter and were cultured in DMEM, FCS 15%, 100 U mL-1 penicillin, 100 μg mL-1 streptomycin. Twenty four h after culture, 10-20 colonies were isolated from each tooth and after 72 h a monolayer was formed.
Immunostaining and FACS analysis of intact mesothelial cells: Adherent cells were trypsinized and the aspirated media was centrifuged at 12000 rpm for 5 min. Cells were washed twice in PBS in 2 mL cryotubues. Cells were blocked with goat serum for 30 min at room temperature. Excess blocker was washed with PBS. Cells were incubated with monoclonal antibodies of conjugated CD34 with FITC diluted to 1/200 and as negative control, CD34 antibody conjugated to nonreactive RPE for 30 min at 4°C. Excess antibody was excluded by washing with PBS. Cells were analyzed by flowcytometry.
Immunocytochemistry (Alkaline phosphatase staining): Cells were fixed with citrate, acetone, formaldehyde fixatives (25/65/8) were prepared in water for 30 sec on coverslips. Having washed the coverslip the cells were exposed to the diazonium salt solution (sodium nitrate, alkaline FRV (1/1) in water plus naphtol As-BL) for 15 min in room temperature. Washed with water jelly tape was used.
RESULTS
Cells from deciduous pulps formed 15-20 colonies in 24 h of culture. After 72 h of culture cells became fully confluent. Figure 1 demonstrates the colony formation (A) and monolayer of deciduous pulp stem cells (B).
As shown in Fig. 2 stem cell colonies from derived from the deciduous pulp showed alkaline phosphatase activity marker. The expression of CD34 is a primitive marker of mesenchyme, which a total percentage of Q2+Q4 = 99/45% of our cells expressed this Marker.
Histogram-1 shows a cell population of similar granular sizes (R1). In graph B the amount of CD 34 expression in Q4 was 88/745 and 5/82% in Q2. Graph C the green colour represents positive cells of approximately 19.94% (R1). Graph D demonstrates the occurrence of the second colour of approximately 1.74% (RN2). Histogram-2 shows a cell population of similar granular sizes (R1). In graph B the amount of CD34 expression in Q4 was 75/68 and 23/77% in Q2. Graph C the green colour represents positive cells of approximately 99/45% (R1). Graph D demonstrates the occurrence of the second colour FL2 of approximately 0% (RN2). Histogram-3 shows a cell population of similar granular sizes (R1). In graph B the amount of CD34 expression in Q4 was 83/36 and 13/45% in Q2 of a total of 96/87%. Graph C the green colour represents positive cells of approximately 95/67 (R1). Graph D demonstrates the occurrence of the second colour of approximately 0/03% (Fig. 3).
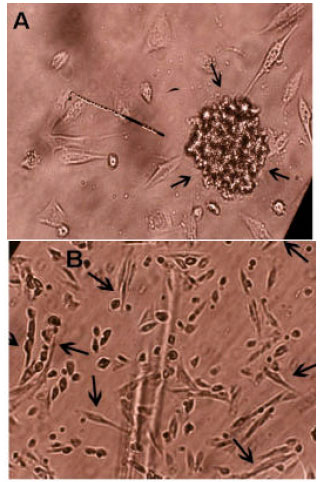 | |
| Fig. 1: | Colony formation of deciduous pulps stem cells: after 24 h of culture stem cells formed colonies (A). The formation of monolayer cells 72 h post culture, dental pulp from a 7 year old female (200x) (B) |
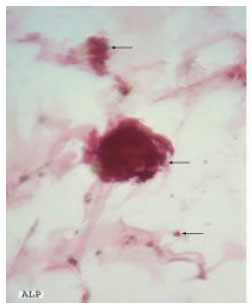 | |
| Fig. 2: | Alkaline phosphatase activity of deciduous pulp stem cells. Accomplishment of immunocytochemistry method revealed positive alkaline phosphatase activity, as reddish stains formed on the colonies are obvious (200X) |
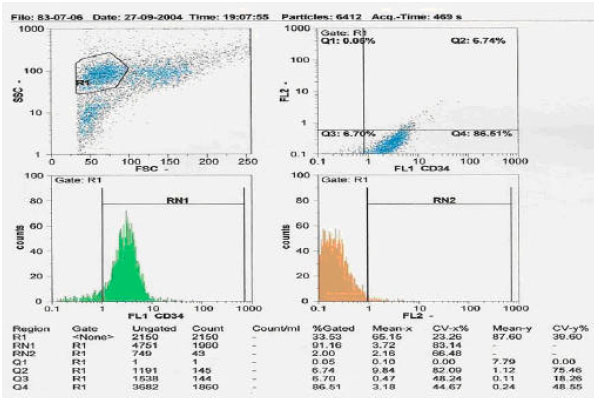 | |
| Fig. 3: | Flowcytometric anlaysis of CD34 expression of stem cells of deciduous pulp: A total percentage of Q2+Q4 = 99/45% of cells expressed CD34 marker |
DISCUSSION
Stem cells are self regenerating, multipotential cells that are capable of producing the full complement of cellular diversity within a region, while progenitor cells are dividing cell that will give rise to a more restricted progeny of a stem cell (Vaccarino, 2000). The isolation of adult stem cells in culture media has been applied in clinical studies. Stem cells from deciduous teeth pulp have also bee studies in previous works, which show a considerable capacity of division and differentiation (Gronthos, 2000; Miura et al., 2003).
In this study pulp cells were isolated by cold trypsin solution treatment. After 24 h colony formation consisting of interdigitating cells was evident. Previous reports have stated that cells from the dental pulp might have an endothelial origin, as they express endothelial specific markers of which CD146 can be mentioned (Grontho et al., 2000; Doyonnas et al., 2001).
Other studies suggest STRO-1 and CD146 expression for these cells as well as differentiation to adentocytes, adiocytes and neurons. From this we could rationally conclude that these cells originate from the neural crest or possibly from the endothelial (Miura et al., 2003). In the present study CD34 marker which is specifically expressed by the progenitors of the endothelial and mesenchyme was studied (Lee et al., 2006; Steidl et al., 2004).
Immunohistochemical analysis reveals the expression of stromal and vascular markers of nestin, alkaline phosphatase and gFGF (Asahara et al., 1997).
The bone marrow consists of three types of stem cells, (a) mesenchymal stem cells, (b) hematopoietic stem cells and (c) endothelial progenitors (Flamme and Risau, 1997).
Mesenchymal stem cells of the bone marrow do not express CD34, in contrast to the other stem cells of that area which significantly express CD34. Hematopoietic stem cells of the bone marrow proliferate in vitro as non adhering cells which form a suspension.
In order to isolate hematopoietic cells BME-2 IGF-1, hEGF-1, hFGF-2, VEGF-1 and ascorbic acid should be added to the media (Peichev et al., 2000), therefore the only appropriate origin of the deciduous stem cells could be the endothelial progenitor cells. Endothelal stem cells have been reported to be mobile in the blood stream.
Having in mind the minute tube connecting to the root of the teeth, an also adding present findings in this study which were the CD34 and alkaline phosphatase activity of these stem cells we propose the probable role of endothelial progenitor cells as well as the neural crest in the derivation of deciduous pulp stem cells.
REFERENCES
- Asahara, T., T. Murohara, A. Sullivan, M. Silver and R. van der Zee et al., 1997. Isolation of putative progenitor endothelial cells for angiogenesis. Science, 275: 964-966.
CrossRefDirect Link - Brazelton, T.R., F.M.V. Rossi, G.I. Keshet and H.M. Blau, 2000. From marrow to brain: Expression of neuronal phenotypes in adult mice. Science, 290: 1775-1779.
Direct Link - D'Amour, K.A. and F.M. Gage, 2002. Are somatic stem cells population or linage vestricted?. Nat. Med., 8: 213-214.
CrossRefDirect Link - Doyonnas, R., D.B. Kershaw and C. Duhme, 2001. Anuria, omphalocele and perinatal lethality in mice lacking the CD34-related protein podocalyxin. J. Exp. Med., 194: 13-27.
Direct Link - Gronthos, S., M. Mankani, J. Brahim, P.G. Robey and S. Shi, 2000. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA., 97: 13625-13630.
CrossRefPubMedDirect Link - Lee, E.S., A.S. Leong and Y.S. Kim, 2006. Calretinin, CD34 and alpha-smooth muscle actin in the identification of peritoneal invasive implants of serous borderline tumors of the ovary. Mod. Pathol., 19: 364-372.
Direct Link - Miura, M., S. Gronthos, M. Zhao, B. Lu, L.W. Fisher, R.P. Gehron and S. Shi, 2003. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA., 100: 5807-5812.
PubMed - Spradling, A. and T.K. Durmmond, 2001. Stem cells find there niche. Nature, 414: 98-104.
PubMedDirect Link - Steidl, U., S. Bork, S. Schaub, O. Selbach and J. Seres et al., 2004. Primary human CD34+ hematopoietic stem and progenitor cells express functionally active receptors of neuromediators. Blood, 104: 81-88.
Direct Link - Vaccarino, F.M., 2000. Stem cell diversity in the CNS. New Scientist., 6: 338-352.
CrossRefDirect Link