
Amal M. Ramadan
National Institute of Oceanography and Fisheries, P.O. Box 182, Suez, Egypt
Magdy M. EL-Halfawy
National Institute of Oceanography and Fisheries, P.O. Box 182, Suez, Egypt
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3120-3125







ABSTRACT
Four economic fish species were collected monthly from commercial catch land at El-Attaka fish landing (Saurida undosquamis, Rhabdosargus haffara, Nemipterus japonicus and Liza carinata). Histoligical examination of ovaries of these species indicated the presence of atretic oocytes (oocyte retention) as a natural phenomenon in fishes. Depending on the histological descriptions atretic oocytes may be classified into two main types: a, bursting b, non bursting. In the present study, frequency of degenerating oocytes were affected by the gonad maturation. In the early stages, the frequency is very low. It increased gradually reaching its maximum in spent stage about 50%. There is a significant difference between the atretic oocytes and gonad maturation (p<0.05). Atresia is also affected by the length of the spawning season (Long or short). All examined ovaries of multiple spawners (S. undosquamis) should atretic oocytes. But in (N. japonicus, R. haffara and L. carinata) about 30 to 35% of the ova show atresia. These species have limited spawning season.
PDF Abstract XML References Citation
How to cite this article
Amal M. Ramadan and Magdy M. EL-Halfawy, 2007. Common Forms of Atresia in the Ovary of Some Red Sea Fishes During Reproductive Cycle. Pakistan Journal of Biological Sciences, 10: 3120-3125.
DOI: 10.3923/pjbs.2007.3120.3125
URL: https://scialert.net/abstract/?doi=pjbs.2007.3120.3125
DOI: 10.3923/pjbs.2007.3120.3125
URL: https://scialert.net/abstract/?doi=pjbs.2007.3120.3125
INTRODUCTION
Atresia is a natural phenomenon and is very common feature of the teleostean ovary. It may be caused by overproduction of oocytes in batch spawners as a result of environmental condition stress (Leino and Maccormik, 1997). Atretic oocytes may be observed in the ovaries amongst the normaloocytes at any stage of development (Simonsen and Gundersen, 2005). The present study aims to describe the different types and common atretic oocytes in the ovary of some red sea fishes during their reproductive cycles and correlate this phenomenon of atresia to gonadal activity of fishes.
MATERIALS AND METHODS
Sample collection: Samples of four Red Sea fish species (S. undosquamis, N. japonicus, R. haffara and L. carinata) were collected from commercial catch at EL-Attaka fish landing. The collection was carried out from September 2003 to May 2004.
Study of reproduction: Samples were dissected and gonads (ovaries) were removed, then weighed. Visual assessment of their maturity stages was made according to Ramadan (2003).
Histological examination: It was carried out by taking a small piece (0.5 cm) from ovary, fixed in Bouin’s solution and transferred to 70% alcohol. The specimens then were dehydrated, cleared and embedded in paraffin wax. Tissue was sectioned by microtome (7 μ) and stained with a heamatoxyline and eosin stains. Microscopic examination of sections was carried out by using a Nikon Microscope.
Statistical analysis: Data were analyzed by using ANOVA , test according to Duncan (1955).
RESULTS
Histological analysis revealed that most of the females of S. undosquamis, N. japonicus, R. haffara and L. carinata were undergoing mass atresia in all developmental stages are evident in oocyte sheath and content. On the basis of histological differences, in Red Sea fishes; atretic follicles were mainly of two types; non-bursting and bursting atresia according to Cupta and Matti (1986).
Non-bursting atresia: This type of atresia is very common in the early oocytes. It is characterized by the non ruptured follicular wall. This type of atresia was further classified into three types:
| • | Capsulated atresia, which is characterized by a drastic reduction in the size of ooplasm which appeared as a dark stain mass (Fig. 1A). |
| • | Lipoidal atresia, in which the follicular wall looked crumpled and nearly thick. The ooplasm loaded with vacuoles which may be lipoid materials (Fig. 2). In late lipoidal atresia the oocyte membrane wrinkled and become thick (Fig. 1B). |
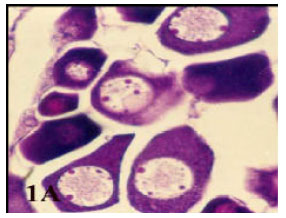 | |
| Fig. 1a: | Photomicrograph of cross section in ovary of Saurida undosquamis showing a non bursting atresia (I-capsulate atresia) appear as a dark stain (Heamatoxylen and Eosin stain X 100) |
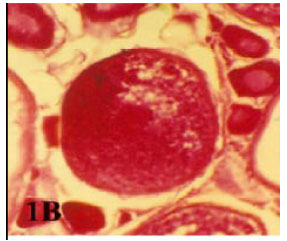 | |
| Fig. 1b: | Photomicrograph of cross section in ovary of Liza carinata showing a non bursting atresia (I-capsulate atresia) appear as a dark stain (Heamatoxylen and Eosin stain X 100) |
| • | Cystic atresia, in which the oocyte lost its normal identity and reduced in size leaving a wide clear pervitelline space between the ooplasm and the oocyte membrane (Fig. 3). |
Bursting atresia: This type was observed in the large developmental oocytes. There were several types of bursting atresia where they identified on the basis of their histological characters.
Multiple bursts: Atretic follicles of these types protrude at several regions of follicles. The wall of oocyte was thicker than normal one (Fig. 4A and B).
Single bursts: In these atretic oocytes the bursting site was single and the contents of the follicle extruded into the stroma. The wall appeared thick (Fig. 5A and B).
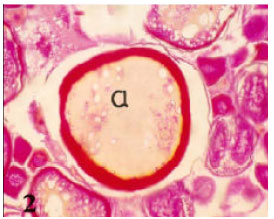 | |
| Fig. 2: | Photomicrograph of cross section in ovary of Nimeptrus japonicus showing a non bursting atresia (II- Lipoidal atresia a) in which the ooplasm loaded with vacuoles (Heamatoxylen and Eosin stain X 50) |
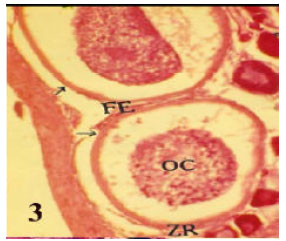 | |
| Fig. 3: | Photomicrograph of cross section in ovary of Rhabdosargus haffara showing a non bursting atresia (III- Cystic) with reduction in size leaving a wide clear pervitelline space. (FE = Follicular Epithelium, ZR = Zona Radiata, OC = Oocyte Content) (Heamatoxylen and Eosin stain X 50) |
Liquified bursts: Atretic follicles of this type were contained large vacuoles in the ooplasm (liquefied) and the oocyte wall was thick and wrinkled in the late stages (Fig. 6A and B and C).
Phagocytic bursts: In this type the follicle cells transformed into phagocytic cells that invade the ooplasm through the ruptured parts of follicular wall. They showed reduction in size and gradual degeneration (Fig. 7).
Finally, all forms of atretic oocytes degenerated and disappeared in the stroma.
Relation between atretic oocytes and ovarian developmental stages of the four red sea fishes: The gonadal status of four fishes (N. japonicus, S. undosquamis, R. haffara and L. carinata) were assessed through the histological examination and the ovarian developmental stages classified, into seven stages, according to Ramadan (2003) as follows, stage I (immature), stage II (developing), stage III (developed), stage IV (fully developed), stage V (gravid), stage VI (spawning) and stage VII (spent) (Fig. 8).
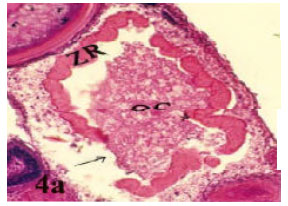 | |
| Fig. 4a: | Photomicrograph of cross section in ovary of Saurida undosquamis showing a bursting atresia (I- multiple bursts) in which the ooplasm loaded with vacuoles (ZR = Zona Radiata, OC = Oocyte Content) (Heamatoxylen and Eosin stain X 50) |
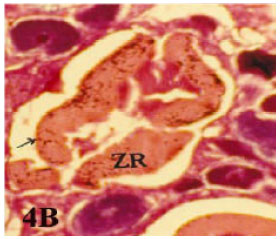 | |
| Fig. 4b: | Photomicrograph of cross section in ovary of Rhabdosargus haffara showing a bursting atresia (I- multiple bursts) in which the ooplasm loaded with vacuoles (ZR = Zona Radiata) (Heamatoxylen and Eosin stain X 50) |
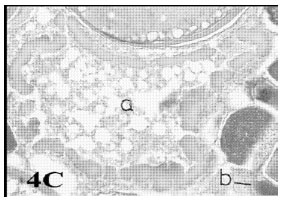 | |
| Fig. 4c: | Photomicrograph of cross section in ovary of N. japonicus showing a bursting atresia (I- multiple bursts) in which the ooplasm loaded with vacuoles (a = oocyte content, b = small cystic type) (Heamatoxylen and Eosin stain X 50). |
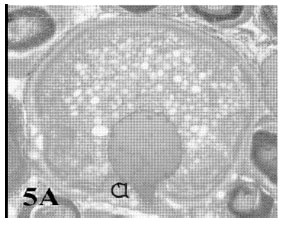 | |
| Fig. 5a: | Photomicrograph of cross section in ovary of Saurida undosquamis showing a bursting atresia (II- single burst a) the content of the oocytes extruded into the stroma (Heamatoxylen and Eosin stain X 50) |
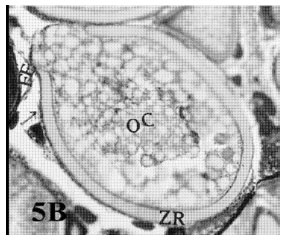 | |
| Fig. 5b: | Photomicrograph of cross section in ovary of Nimeptrus japonicus showing a bursting atresia (II- single burst) the content of the oocytes extruded into the stroma (FE = Follicular Epithelium, ZR = Zona Radiata, OC = Oocyte Content) (Heamatoxylen and Eosin stain X 50) |
Examination of gonads revealed that, fishes were in immature phase (stage I) no atretic oocytes were found.
However, in stage II according the above scheme contained a very small percent of atretic oocytes to normal ones (1 and 2%, respectively).
Evidently, in the mature phase (from stage III to stage VII) the percentage was increase reaching its maximum values in the spawning(stage VI) and spent (stage VII) stages (15 and 50%, respectively).
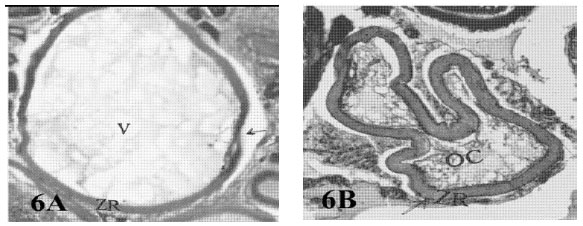 | |
| Fig. 6a: | Photomicrograph of cross section in ovary of Liza carinata showing a bursting atresia (III-liquefied burst) (V = Vacuoles, ZR = Zona Radiate) (Heamatoxylen and Eosin stain X 50) |
| b: | Photomicrograph of cross section in ovary of Rhabdosargus haffara with a bursting atresia showing late liquefied with wrinkled and thick oocyte membrane (ZR = Zona Radiata, OC = Oocyte Content) (Heamatoxylen and Eosin stain X 50) |
| Table 1: | Frequency of ovarian atresia in four Red Sea fish species (September 2003 to May 2004) |
 | |
| aRammadan (1995), bEL-Halfawy (1995), cEL-Halfawy (2001) and dEL-Halfawy (2004) | |
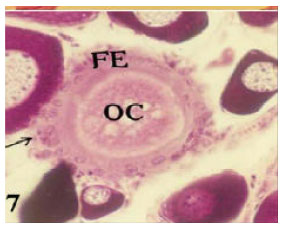 | |
| Fig. 7: | Photomicrograph of cross section in ovary of Saurida undosquamis showing a bursting atresia (IV-phagocytic bursts) (FE = Follicular Epithelium, OC = Oocyte Content) (Heamatoxylen and Eosin stain X 50) |
Atresia and spawning season: From Table 1 frequency of ovarian atresia was marked higher in fishes which characterized by long spawning season like S. undosquamis than in short spawning season like others species (N. japonicus, R. haffara and L. carinata). In S. undosquamis (amultiple spawner) in which spawning season extend allover the year (Ramadan, 1995) all the examined females show atretic follicles (100%).
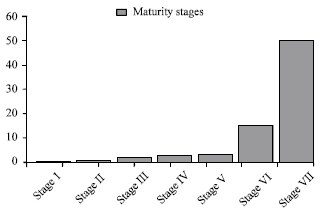 | |
| Fig. 8: | Frequency of atretic oocytes in relation to ovarian developmental stages of some Red Sea fishes (2003) |
Meanwhile, in the partial spawning fishes in which spawning extends about 3 to 4 months in the year, the frequency of atretic females ranged from 30% in L. carinata to 35% in N. japonicus and R. haffara.
DISCUSSION
Several factors have been associated with egg retention by female fishes including overcrowding, environmental activity, elevated concentration of heavy metal and spawning success (Levavi-Sivan et al., 2004). Degeneration of oocytes has been observed in the ovaries of all teleosts that have been carefully examined histologically (Tripple and Harvey, 1990). These atretic follicles may be a source of progestational hormone, they appear to function chiefly in the production of nutritive fluid (Wallace and Selman, 1981). Atresia occurs when oocytes abort to develop and fail to be spawned from the ovary and then are resorbed back into the gonad (Arocha, 2002).
Study of serial sections of the ovary of four Red Sea fish species (S. undosquamis, N. japonicus, R. haffara and L. carinata) revealed two main types of atresia i.e., nonbursting and burusting described as a common types. Ramadan et al. (1987) indicated that deformation of the wall of oocyte was considered as the first step of atresia and phagocytosis of oocytes. Kamel (1990) classified the atretic follicles according to the diameters, but in the present study classification depended on the histological descriptions. As in the finding of Ramadan et al. (1987), the atresia for both types nonbursting and bursting begin from 2 to 15% of oocytes fail to undergo maturation or ovulation in the peri and during spawning in stages (II to VI stages) and about 50% in the post spawning (in stage VII) spent stage.
There was a significant difference in maturity level with atretic oocytes (p<0.05) and increased significantly in stage VII (p<0.02). So, in the post spawning stage the atresia become most abundant immediately after egg laying. These findings were confirmed with Lowerre- Barbier et al. (1996) who reported that the atretic follicles occur at any stage of follicular development. Higher prevalence of atresia in the early vitellogenic phase has been described as a natural process regulating the surplus of oocytes in the early vitellogenic stage recruited into successive stage of development (Bromley et al., 2000). However, Erpino (1973) and Assem and EL-Zaeem (2005) stated that atresia does not occure in the pre-vitellogenic developmental stages. Similar suggestion have been reported by Trippel and Harvey (1990). There was no evidence that atresia interfered with gamete development subsequently.
In the present study, the ratio of atresia increased by gonad maturation reaching its maximum values in the spent stage VII and this results agree with results obtained by Kurita et al. ( 2003). They stated that atresia has been noted to be wide spread in later phase of maturation process and associated with the a valuable energy resources and environmental conditions. But this disagreement with Simonsen and Gundersen (2005) who reported that atresia was highest in early phases of maturation in Green land halibut but relatively high levels of atresia were also observed in fish in more advanced maturity phase.
However the investigators did not correlate atresia to spawning season and fish activity. Our study touch this correlation when investigate four species differ in the time and long of spawning season. As far as known, the multiple spawners as S. undosquamis, which have repetitive cycles of spawning when spawn of one brood the oocytes of the next brood are maturing. The atretic ovaries found allover the year and in all gonadal maturity by 100% of examined ovaries. This may be due to the higher activity of this species and the over production causes in failer of some oocytes to be spawned.
On the other hand, the other species which have a limited or short spawning season like N. japonicus, R. haffara and L. carinata, the atretic ovaries occurred in about 3 to 30% of examined females. This ratio concentrated after spawning season and few ratio in peri and during spawning season was observed. This also may be due to its limit activity in egg production. So, atresia may be important as it affects the estimated spawning potential of stock. Besides the previous study and earlier workers on atresia phenomenon, further investigation to understand the mechanism and significance of atresia to fish behavior.
REFERENCES
- Arocha, F., 2002. Oocyte development and maturity classification of swordfish from the North-Western Atlantic. J. Fish Biol., 60: 13-27.
CrossRefDirect Link - Assem, S.S. and S.Y. El-Zaeem, 2005. Application of biotechnology in fish breeding. II: Production of highly immune genetically modified redbelly tilapia, Tilapia zillii. Afr. J. Biotechnol., 4: 449-459.
Direct Link - Fair, T., 2003. Follicular oocyte growth and acquisition of developmental competence. Anim. Reprod. Sci., 78: 203-216.
CrossRef - Levavi-Sivan, B., Viman, O. Sachs and A. Tzchori, 2004. Spawning induction and hormonal levels during final oocyte maturation in the silver perch. Aquacult, 229: 419-431.
CrossRefDirect Link