
Zohreh Hojati
Department of Biology, Faculty of Sciences, Isfahan University, Isfahan, Islamic Republic of Iran
Majid Motovali-Bashi
Department of Biology, Faculty of Sciences, Isfahan University, Isfahan, Islamic Republic of Iran
Naser Golbang
Department of Biology, Faculty of Sciences, Isfahan University, Isfahan, Islamic Republic of Iran
Farshad Darvishi
Department of Biology, Faculty of Sciences, Isfahan University, Isfahan, Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 18 | Page No.: 3079-3084







ABSTRACT
The genetics of streptomycin production is well characterized in Streptomyces griseus. More than 25 clustered genes encode proteins involved in biosynthesis, regulation and transport functions. StrR, the pathway specific transcriptional activator or regulator that located in this cluster, then induces transcription of other streptomycin production genes by binding multiple sites in the gene cluster. We aim to put strR gene in to different multicopy and integrated expression vector specifically designed for Streptomyces. To start with, the isolated strR gene was ligated into pBluescript (pBs) vector and transformed into different strains of Escherichia coli. The correct structure of the recombinant plasmid, isolated from transformed E. coli, was confirmed using gel electrophoresis, PCR and double digested with restriction enzymes BamHI and EcoRI. Finally the plasmid map, named pFDstrR. This unique vector has a much expanded Multiple Cloning Site (MCS), which makes it suitable for different purposes of gene cloning and also site directed mutagenesis or gene targeting. This gene will be lifted up and transfer into different varieties of Streptomyces specific vectors in order to make different transgenic or genetically manipulated Streptomyces.
PDF Abstract XML References Citation
How to cite this article
Zohreh Hojati, Majid Motovali-Bashi, Naser Golbang and Farshad Darvishi, 2007. Detection of Cloned strR, an Antibiotic Regulatory Gene, using RFLP and Nested PCR. Pakistan Journal of Biological Sciences, 10: 3079-3084.
DOI: 10.3923/pjbs.2007.3079.3084
URL: https://scialert.net/abstract/?doi=pjbs.2007.3079.3084
DOI: 10.3923/pjbs.2007.3079.3084
URL: https://scialert.net/abstract/?doi=pjbs.2007.3079.3084
INTRODUCTION
Streptomyces species are mycelial, aerobic gram-positive bacteria that are readily isolated from soil (Kim et al., 2005; Horinouchi et al., 2001). Streptomyces are unique amongst prokaryotes due to their complicated morphological differentiation (Anderson et al., 2002). These morphological changes are accompanied by a wide range of physiological events including the production of secondary metabolites, many of which have potentially important biological activities. They include many useful antibiotics and other products such as the antitumor drugs and herbicide too (Aigle et al., 2005; Hesketh et al., 2001).
Streptomyces griseus produces the antibiotic streptomycin and forms spores even in liquid culture (Babcock and Kendrick, 1988). In Streptomyces griseus, an autoregulatory factor called A- factor that act as a bacterial hormone, positively regulates both physiological and morphological differentiation (streptomycin production and spore formation) at a concentration as low as 10-9 M (Yamazaki et al., 2000). It has been elucidated that A-factor acts as a switch for physiological and morphological differentiation messenger in signal transduction in eukaryotes, as well as in catabolite repression in Gram-negative bacteria (Ohnishi et al., 2005; Horinouchi et al., 2001).
The first of the aminoglycoside antibiotics, streptomycin, was reported in 1944, Streptomycin proved to be effective for the first time in the treatment of tuberculosis (Ramón-García et al., 2006; Schatz et al., 1944). Streptomycin and related compounds (SARCs) are three-ringed structures principally derived from glucose-6-phosphate by a complex, branched biosynthesis pathway (Anderson et al., 2002). Streptomycin is one of the best-studied antibiotics at the biochemical and genetic levels. The genetics of streptomycin production is well characterized in Streptomyces griseus N2-3-11, for which more than 25 clustered genes have been, described (Fig. 1) that encode biosynthesis, regulatory and transport functions (Egan et al., 1998).
The A-factor autoregulator induces the transcription of adpA, the central positive regulator, by inactivating ArpA, which acts as a transcriptional repressor for adpA (Yamazaki et al., 2003a,b; Ohnishi et al., 1999). AdpA then pleiotropically induces the transcription of genes, such as adsA and strR, encoding specific positive transcriptional regulatory proteins for morphogenesis and streptomycin biosynthesis, respectively (Hong et al., 2007).
The strR gene encoding a pathway-specific regulator, by binding an upstream activation sequence, about 270 bp upstream from the transcriptional start point of strR (Vujaklija et al., 1993). The pathway specific transcriptional activator, strR, then induces transcription of other streptomycin production genes by binding multiple sites in the gene cluster, leading to biosynthesis of streptomycin from glucose (Browning and Busby, 2004; Retzlaff and Distler, 1995).
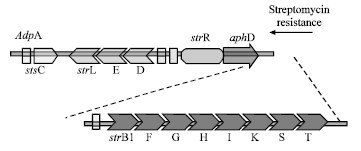 | |
| Fig. 1: | Some of the almost identified genes in Streptomycin biosynthesis gene cluster: strR: Cluster regulatory gene; strA (aphD): phosphotransferase (streptomycin resistance); promoters are shown in oblongs; direction of the genes are illustrated by arrows. |
The main goal in this research is production of elevated levels of streptomycin, using transgenic Streptomyces. To gain this, we initially need the regulatory gene for streptomycin production, strR. This gene was successfully isolated from Streptomyces griseus and transformed into Escherichia coli (Darvishi et al., 2006). The correct structure of the gene and recombinant vector was then confirmed in this research, using Nested PCR and RFLP-PCR.
MATERIALS AND METHODS
This study was conducted in Genetic Research Laboratory at Isfahan University during years 2005-2006.
Bacterial strains: In this study Streptomyces griseus (PTCC 1172: from Iranian Industrial and Science Research Institute) and different strains of Escherichia coli (including DH5α, XL1-Blue and HB101, from Sinagen Iran) were used. The Streptomyces strain was grown on standard liquid and agar media at 30°C and stored as spore suspensions in 20% glycerol at -20°C (Kieser et al., 2000). Luria-Bertani (LB) medium and agar was used for propagation of E. coli at 37°C, for both solid and liquid media and stored 20% glycerol at -70°C.
Vector: The pBluescript SK reproduced from Stratagene Catalogue used as Vector in this study.
Primers: All primers used in this investigation listed in Table 1. The computer program OLIGO (version 5.0, W. Rychlik) was used to design and check primers. Two sets of primers were designed to not only amplify the strR region but also integrate one unique recognition sites (BamHI and XbaI) in each end of the amplified fragment.
Isolation of total DNA from Streptomyces: Usage of liquid culture to isolate total DNA from Streptomyces. Isolation of total DNA from Streptomyces was carried out using the High Pure PCR Template Preparation Kit (Roche; Cat. No. 1 796 828). The amount of DNA was quantified by gel electrophoresis and then spectrophotometer analysis.
PCR: The reaction mixture for PCR amplification was prepared as follows: primer forward, 20 ng; primer reverse, 20 ng; dimethyl sulfoxide, 4 μL; 10X PCR buffer with MgSO4, 5 μL; deoxynucleoside triphosphates (dNTPs), 2 μL (10 mM each dNTP); H2O, to 50 μL. DNA was added to the reaction mixture, 100 ng of chromosomal DNA as a template. The PCR cycling protocol used a hot start of 94°C for 10 min, after which the temperature was reduced to 80°C to allow the addition of 0.3 μL (2.5 U) of Pfu polymerase and Cycling then continued with a denaturation temperature of 94°C for 45 sec, an annealing temperature of either 66°C (strR1) or 62°C (strR2 and strR3) for 30 sec, an extension temperature of 72°C for 1 min. The number of cycles carried out was normally 25. After amplification, the product was visualized by standard electrophoresis procedures with 0.7% (wt/vol) agarose gels and TBE buffer.
Restriction Endonuclease (RE) digestion: Digestion was performed following the recommendations of suppliers (Fermentas). DNA samples such as plasmid or chromosomal DNA (0.2-5 μg) were generally digested with 5-10 units of restriction enzyme (BamHI and XbaI) in a 10-20 μL final volume of restriction buffer (10 x buffer) for about 1-3 h in a water bath at the recommended temperature (normally 37°C). A sample was run on an agarose gel after incubation with each enzyme to ensure complete digestion (Hojati, 2002).
| Table 1: | Different sets of primers designed and used in this study |
 | |
DNA ligation: DNA ligation normally was done using 1 unit of T4 DNA ligase in the presence of 1 x ligation buffer. To optimize the ligation results, the amount of insert DNA was three times that of the vector DNA and incubation was done either at 16°C overnight (Hojati, 2002).
Transformation of E. coli: For making competent cells from E. coli, the calcium chloride method was used (Hojati, 2002). Prior to transformation the frozen competent cells (200 μL aliquot) were slowly thawed on ice for about 30 min. The DNA to be transformed was added, 50 ng of pBluescript SK plasmid preparation. After mixing gently and incubation on ice for 30 min, the cells were heat shocked at 42°C for 90 sec. Heat shocked cells were added to 2 mL LB (in absence of any antibiotic) and incubated at 37°C with shaking for 1 h. At this stage cells were ready for inoculation on to LB plates, a 100 μL sample was spread onto the LB plate containing antibiotic selection. The plates were allowed to dry before incubating overnight at 37°C (Hojati, 2002).
Extraction of plasmid DNA from E. coli: The E. coli DNA was isolated according to the method described by Holmes and Quigley (Hojati, 2002).
RESULTS
Total DNA was isolated from Streptomyces and then was subjected to gel electrophoresis to analyze the concentration and purities. The isolated total DNA was entirely pure and very condenses. This DNA was used for PCR reactions. Three different sets of primer were used for different purposes. One set of primers (strR1) were designed (using OLIGO5) to identify and isolate strR from the genome. Other primers (strR2 and strR3) were designed and then subjected to modification at each 5' end, in order to have recognition sites for BamHI and XbaI sites. These primers then not only amplify the strR gene but also integrate one unique recognition site in each end of the amplified fragments (Fig. 2). The PCR condition was set up for the modified primers and the resultant fragments were gel electrophoresed. Two different sets were successfully amplified strR gene (Fig. 3). The isolated fragment has to be confirmed. Two different strategies were conducted to confirm the amplified fragment as strR gene. Initially, the first set of primer (strR1) was again used to amplify the strR PCR product (Fig. 3). So the performance of Nested PCR, using this set of primer was clearly confirmed the existence of strR gene (Fig. 3).
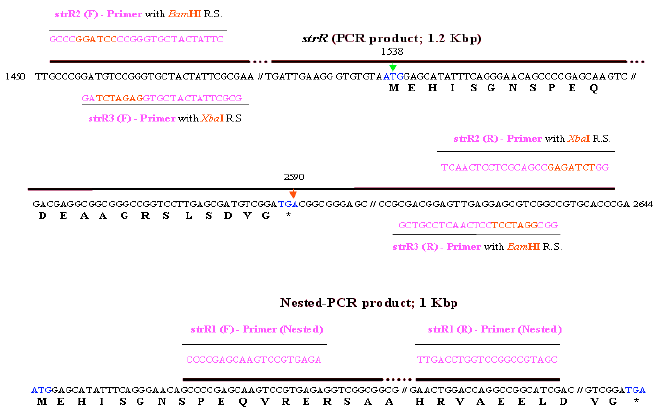 | |
| Fig. 2: | The nucleotide sequence of strR. Primers are shown by bold lines, with the name of each primer on each line. The size of each amplified fragment is indicated in kbp (Darvishi, 2005) |
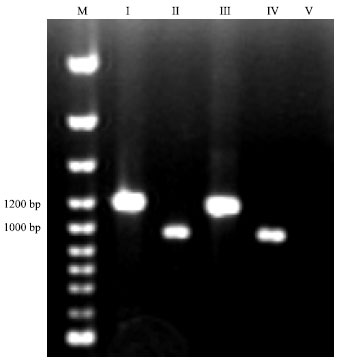 | |
| Fig. 3: | A: PCR amplified fragments, strR, amplified and Confirmation of them using the amplification of PCR products by Internal primers, strR1 (forward and reverse) were used in a PCR cycle named nested PCR, which visualized by gel electrophoresis (0.7%). I: The strR amplified with strR2 primer set and Pfu polymerase. II: PCR product of strR2 primer set amplified by strR1 primer set. III: The strR amplified with strR3 primer set and Pfu polymerase. IV: PCR product of strR3 primer set amplified by strR1 primer set. V: Negative control. M: Indicates 100 bp DNA ladder markers |
PCR-RFLP then was carried out using two different enzymes AluI and BglII. AluI cuts the strR at 665 and 1037, producing three fragments, 665, 372 and 152 bp. The second enzyme, BglII, does not cut the strR. The resultant fragments were confirmed the correct fragment for the strR gene (Fig. 4). Finally, although the Pfu polymerase has got a unique proofreading feature, that do not normally leave any mutation, but still the entire fragment has to be sequenced to make sure that the polymerase did not make any mistake; this step is currently in progress.
strR cloning: at this step, the strR fragment is ready for transformation. E. coli DH5α, then was transformed with pBs plasmid and the transformed strains were initially confirmed using antibiotic selection. pBs was isolated from the transformed E. coli. The PCR amplified fragment was then double digested with BamHI and XbaI and the resultant fragment was subjected to gel purification. The vector, pBs plasmid, also was double digested and gel purified. A ligation mixture was set up using double digested vector and strR.
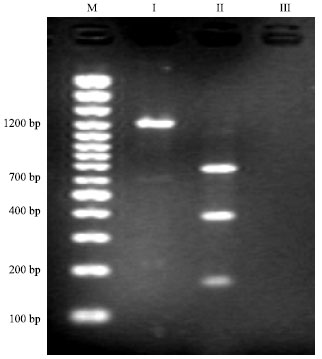 | |
| Fig. 4: | Identification of the amplified strR, using restriction digestion analysis named as RFLP-PCR. I: The BglII enzyme does not cut the strR Fragment. II: AluI enzyme cuts the strR at 665 and 1037, producing three fragments, 665, 372 and 152 bp. III: Negative control. M: Indicates 100 bp DNA ladder marker. The results visualized by gel electrophoresis (2%). The resultant fragments were confirmed the correct fragment for the strR gene |
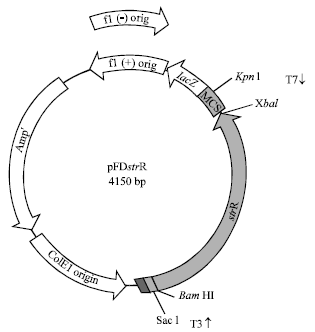 | |
| Fig. 5: | The physical map of plasmid pFDstrR, 4.1 kb, a derivative of pBluescript plasmid containing strR gene. Not all the restriction sites are drawn |
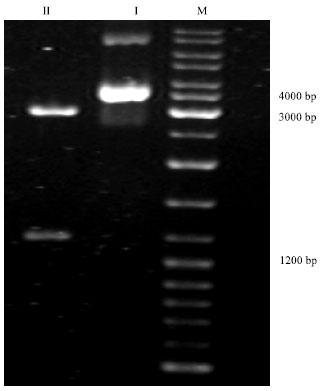 | |
| Fig. 6: | Identification of the pFDstrR plasmid using gel electrophoresis and cut with BamHI and XbaI. I: The pFDstrR plasmid. II: The pFDstrR plasmid cut with BamHI and XbaI. M: Indicates Gene Ruler™ DNA Ladder Mix DNA marker. The plasmid preparation was subjected to gel electrophoresis (0.7%) |
Competent cells of E. coli DH5α and E. coli XL1-Blue were transformed, using 10 μL of ligation mixture. The transformation efficiencies, for both strains, were quite high, so there was a bout 10-20 colonies on the plate that was inoculated with 100 μL of the transformed cells of E. coli XL1-Blue.
Almost the same result was observed with the E. coli HB101. Plasmid preparation was carried out on transformed colonies to identify the structure of the recombinant plasmids; pFDstrR, 4.1 kb, Figure 5. The isolated plasmid was gel electrophoresis to identify the right size and then cut with BamHI and XbaI (Fig. 6). The pFDstrR was then used as PCR template in a PCR reaction with a set of primer that could confirm the existence of strR. The correct recombinant plasmid, therefore do exist in the new strain of E. coli.
DISCUSSION
The overall aim of this study was to further our knowledge of the regulation of antibiotic production in Streptomyces (the producer of two thirds of all known microbial antibiotics) and to manipulate production of antibiotics, especially with the view to designing novel antibiotics with desired activities. In our previous work, it's been reported that cdaR, regulatory gene for the production of Calcium Dependent Antibiotic (CDA), autoregulates its own transcription. Therefore introducing extra copies of cdaR into different strains of Streptomyces coelicolor MT1110, S. coelicolor 2377 and Streptomyces lividans led to an increase in the CDA and also overproduction of this antibiotic (Hojati, 2002). Designing novel antibiotics, on the other hand, greatly dependent on the structural analysis of the gene cluster for each antibiotic (Hojati et al., 2002). Streptomycin, an aminoglycoside antibiotic, proved to be effective for the first time in the treatment of tuberculosis (Ramón-García et al., 2006; Schatz et al., 1944). Streptomycin and related compounds (SARCs) are three-ringed structures principally derived from glucose-6-phosphate by a complex, branched biosynthesis pathway (Anderson et al., 2002).
This study started at the point where a regulatory gene, strR, for streptomycin production had been identified. The strR initially was isolated from Streptomyces griseus, one of the major producers of Streptomycin, using a set of designed primer. The isolated fragment was confirmed as strR gene using a set of designed internal primers in a technique described as Nested PCR. We aim to put this gene in to different multicopy and integrated expression vector specifically designed for Streptomyces. To start with, the strR gene was ligated into pBluescript (pBs) and transformed into different strains of E. coli (including DH5α, XL1-Blue and HB101). The correct structure of the recombinant plasmid, isolated from transformed E. coli, was confirmed using gel electrophoresis, PCR and double digested with BamHI and XbaI. Finally the plasmid map, named pFDstrR, was drawn using Plasmid software (Fig. 5).
This unique vector has got a much expanded Multiple Cloning Site (MCS), which makes it suitable for different purposes of gene cloning. This gene will be lifted up and transfer into different varieties of Streptomyces specific vectors in order to make different transgenic Streptomyces. These strains hopefully are able to produce elevated levels of Streptomycin in comparison with the original strain.
ACKNOWLEDGMENT
This work was supported by the Isfahan University Graduate office and Research office (Grant No. 831021).
REFERENCES
- Aigle, B., A. Wietzorrek, E. Takano and M.J. Bibb, 2005. Involvement of AlpV, a new member of the Streptomyces antibiotic regulatory protein family, in regulation of the duplicated type II polyketide synthase alp gene cluster in Streptomyces ambofaciens. J. Bacteriol., 187: 2491-2500.
CrossRefDirect Link - Browning, D.F. and S.J. Busby, 2004. The regulation of bacterial transcription initiation. Nat. Rev. Microbiol., 2: 57-65.
CrossRefDirect Link - Egan, S., P. Wiener, D. Kallifidas and E.M.H. Wellington, 1998. Transfer of streptomycin biosynthesis gene clusters within Streptomycetes isolated from soil. Applied Environ. Microbial., 64: 5061-5063.
PubMedDirect Link - Hesketh, A., J. Sun and M. Bibb, 2001. Induction of ppGpp synthesis in Streptomyces coelicolor A3(2) grown under conditions of nutritional sufficiency elicits actII-ORF4 transcription and actinorhodin biosynthesis. Mol. Microbiol., 39: 136-144.
PubMedDirect Link - Hojati, Z., C. Milne, B. Harvey, L. Gordon and M. Borg et al., 2002. Structure, biosynthesis origin and engineered biosynthesis of calcidium-dependent antibiotics from Streptomyces coelicolor. Chem. Biol., 9: 1175-1187.
PubMedDirect Link - Hong, B., S. Phornphisutthimas, E. Tilley, S. Baumberg and J.K. McDowall, 2007. Streptomycin production by Streptomyces griseus can be modulated by a mechanism not associated with change in the adpA component of the A-factor cascade. Biotechnol. Lett., 29: 57-64.
PubMedDirect Link - Horinouchi, S., Y. Ohnishi and D.K. Kang, 2001. The A-factor regulatory cascade and cAMP in the regulation of physiological and morphological development in Streptomyces griseus. J. Indust. Microbiol. Biotechnol., 27: 177-182.
CrossRefDirect Link - Kim, D.W., K.F. Chater, K.J. Lee and A. Hesketh, 2005. Effects of growth phase and the developmentally significant bldA-specified tRNA on the membrane-associated proteome of Streptomyces coelicolor. Microbiology, 151: 2707-2720.
CrossRefDirect Link - Ohnishi, Y., S. Kameyama, H. Onaka and S. Horinouchi, 1999. The A-factor regulatory cascade leading to streptomycin biosynthesis in Streptomyces griseus: Identification of a target gene of the A-factor receptor. Mol. Microbiol., 34: 102-111.
PubMedDirect Link - Ohnishi, Y., H. Yamazaki, J.Y. Kato, A. Tomono and S. Horinouchi, 2005. AdpA, a central transcriptional regulator in the A-factor regulatory cascade that leads to morphological development and secondary metabolism in Streptomyces griseus. Biosci. Biotechnol. Biochem., 69: 431-439.
Direct Link - Ram�n-Garc�a, S. I. Otal, C. Mart�n, R. G�mez-Lus and J.A. A�nsa, 2006. Novel streptomycin resistance gene from Mycobacterium fortuitum. Antimicrob. Agents Chemother., 50: 3920-3922.
CrossRefPubMedDirect Link - Yamazaki, H., Y. Ohnishi and S. Horinouchi, 2000. An A-factor-dependent extracytoplasmic function sigma factor (sigma adsA) that is essential for morphological development in Streptomyces griseus. J. Bacteriol., 182: 4596-4605.
PubMedDirect Link - Yamazaki, H., Y. Ohnishi and S. Horinouchi, 2003. Transcriptional switch on of ssgA by A-factor, which is essential for spore septum formation in Streptomyces griseus. J. Bacteriol., 185: 1273-1283.
CrossRefDirect Link - Yamazaki, H., Y. Takano, Y. Ohnishi and S. Horinouchi, 2003. AmfR, an essential gene for aerial mycelium formation, is a member of the adpA regulon in the A-factor regulatory cascade in Streptomyces griseus. Mol. Microbiol., 50: 1173-1187.
PubMedDirect Link