
M. Farkhari
Department of Agronomy and Plant Breeding, Agricultural College, University of Tehran, Karaj, Iran
M. R. Naghavi
Department of Agronomy and Plant Breeding, Agricultural College, University of Tehran, Karaj, Iran
S. A. Pyghambari
Department of Agronomy and Plant Breeding, Agricultural College, University of Tehran, Karaj, Iran
Sabokdast
Department of Agronomy and Plant Breeding, Agricultural College, University of Tehran, Karaj, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 17 | Page No.: 2868-2873







ABSTRACT
Genetic variation of 28 populations of jointed goatgrass (Aegilops cylindrica Host.), collected from different parts of Iran, were evaluated using both RAPD-PCR and SDS-PAGE of seed proteins. The diversity within and between populations for the three-band High Molecular Weight (HMW) subunits of glutenin pattern were extremely low. Out of 15 screened primers of RAPD, 14 primers generated 133 reproducible fragments which among them 92 fragments were polymorphic (69%). Genetic similarity calculated from the RAPD data ranged from 0.64 to 0.98. A dendrogram was prepared on the basis of a similarity matrix using the UPGMA algorithm and separated the 28 populations into two groups. Confusion can happen between populations with the same origin as well as between populations of very diverse geographical origins. Our results show that compare to seed storage protein, RAPD is suitable for genetic diversity assessment in Ae. cylindrica populations.
PDF Abstract XML References Citation
How to cite this article
M. Farkhari, M. R. Naghavi, S. A. Pyghambari and Sabokdast, 2007. Genetic Variation of Jointed Goatgrass (Aegilops cylindrica Host.) From Iran Using RAPD-PCR and SDS-PAGE of Seed Proteins. Pakistan Journal of Biological Sciences, 10: 2868-2873.
DOI: 10.3923/pjbs.2007.2868.2873
URL: https://scialert.net/abstract/?doi=pjbs.2007.2868.2873
DOI: 10.3923/pjbs.2007.2868.2873
URL: https://scialert.net/abstract/?doi=pjbs.2007.2868.2873
INTRODUCTION
Genetic resources are the gene pool available for breeders and other scientists, and in the Triticeae tribe several pools are recognized (Von Botmer et al., 1992; Gill et al., 2006). Aegilops is characterized as a Mediterranean-Western Asiatic element and its center of diversity follows the central part of the Fertile Crescent arc in West Asia (Hegde et al., 2002).
Ae. cylindrica Host. (genome formula: CCDD) is an amphiploid, resulting from hybridization between the diploids Ae. caudata (CC) and Ae. squarrosa (DD). This species is, however, frequent in the Mediterranean area and in the Middle East, which is its center of distribution (Guadagnuolo et al., 2001). This species distributes of frequently along roadsides, edges of cultivation, dry hillsides, and grassy steppes, where it tolerates disturbance (van Slageren, 1994). It is known to hybridize spontaneously with wheat (Gandilyan and Jaaska, 1980; Snyder et al., 2000). Naturally formed hybrids between Ae. cylindrica and wheat occasionally produce seeds, presumably via spontaneous cross-pollination, which could facilitate gene flow between the D genomes of the two species (Gandilyan and Jaaska, 1980; Guadagnuolo et al., 2001; Galaev and Sivolap, 2005). Therefore, understanding the genetic structure of these troublesome weedy populations may facilitate designing new ways to control them, especially when most other conventional methods of weed control have failed to contain their spread (Donald and Ogg, 1991).
Today, molecular tools are available to evaluate and characterize, to a certain extent, genetic diversity among gene bank accessions. Genetic diversity of Ae. cylindrica has been evaluated using morphological data (Dotray and Young, 1993), isozyme analysis (Watanabe et al., 1994; Hegde et al., 2002), protein variation (Wan et al., 2002) and molecular markers (Okuno et al., 1998; Guadagnuolo et al., 2001; Pester et al., 2003). But although many landraces of wheats were collected in Iran and were available before and after the Islamic Revolution, wild wheat and Aegilops from Iran are still largely unknown. The objective of this study was to understand the extent and pattern of genetic diversity in 28 populations of allotetraploid wild wheat Ae. cylindrica, collected from roadside of Iran, using seed storage protein and RAPD markers.
MATERIALS AND METHODS
Plant material: Twenty eight populations of Aegilops cylindrica which collected from origin sites of Iran in 2004, including 10 provinces, (Table 1) were used in this study.
Storage protein analysis: In order to evaluate genetic diversity of high molecular weight subunit of glutenin within and between populations, three samples were taken and analysed from each of 28 populations of Ae. cylindrica. Chinies spring cultivar was used as a standard to compare subunit composition. The seed were crushed after removal of the embryo.
| Table 1: | Abbreviation, origin site and province of populations of Ae. cylindrica analyzed |
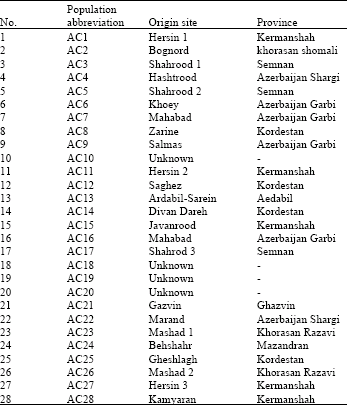 | |
The flour was mixed in an extraction buffer of 62 mM Tris-HCL (pH = 6.8) buffer containing 10% (W/V) glycerol, 4% (W/V) Sodium Dodecyl Sulfate (SDS), 0.01% (W/V) bromophenol blue and 4% ß-mercaptoethanol. Samples were allowed to stand at room temperature for at least 12 h with occasional vortexing. They were then placed in a boiling waterbath for 2 min and then centrifuged for 5 min at 6500 rpm and 15 μL of each sample was loaded on the gel. Proteins were fractionated by SDS-PAGE according modified of Laemmli (1970) procedure using stacking gel Proteins were fractionated using stacking gel containing 4% acrylamide, 0.05% bis acrylamide, 0.1% SDS and 0.00009 M Tris-HCL and separating gel containing 10% acrylamide, 0.13% bis acrylamide, 0.1% SDS and 6.78 M Tris-HCL. Gels were stained overnight with 0.01% (W/V) Coomassie Brilliant Blue R 250 in water and acetic acid (10%) and then destained overnight in water for at least 24 h.
RAPD assay: Total genomic DNA was isolated from young leaves of greenhouse-grown plants according to the CTAB protocol (Saghai-Maroof et al., 1984) with minor modifications. To reveal the level of genetic variation for each population, DNA of five plants were bulked and analysed. A total of fifteen 10-mer oligonucleotides from UBC (University of British Colombia) series were selected according to the number and consistency of amplified fragments. PCR reactions were carried out in a 25 μL-1 volume containing 1 unit of Taq polymerase, 5 ng of genomic DNA template, 0.2 μmol-1 of primer, 2 m μ of each dATP, dCTP, dGTP and dTTP, and 2.5 μL of 10X PCR reaction buffer. Amplifications were performed in a DNA Thermo-cycler (Perkin-Elmer) programmed for 45 consecutive cycles each consisting of 1 min at 92°C, 1 min at 37°C and 2 min at 72°C. Following amplification, the samples were subjected to electrophoresis in 6% acrylamide gels in TAE buffer running at 200 v for 2.5 h. The gels were stained using etidum bromide and viewed under Ultra-violet light.
Data analysis: Polymorphic RAPD fragments were scored as either present (1) or absent (0) across all populations. Only distinct, well-resolved fragments were scored. Binary matrix was used to estimate the genetic similarities between pairs, by employing Dice index (Nei and Li, 1979). These similarity coefficients were used to construct dendrogram using the Unweighted Pair Group Method with Arithmetic averages (UPGMA) employing the SAHN (Sequential, Agglomerative, Hierarchical, and Nested clustering) from the NTSYS-PC (Numerical Taxonomy and Multivariate Analysis System), version 2.02 (Applied Biostatistics) program (Rohlf, 1998). A principal coordinate analysis (PCO) was also conducted with the same program using the DCENTER and EIGEN procedures.
RESULTS
Protein analysis: The patterns of high molecular weight (HMW) glutenin seed proteins among 28 populations of jointed goatgrass detected that diversity in the three-band HMW glutenin pattern was extremely low. In other words, there was very low diversity within and between studied populations. In previous study, Wan et al. (2002) identified 3 subunits bands as 1Cx, 1Cy and 1Dy in all accessions of Ae. cylindrica. In this study, we also found 3 subunits bands for all within and between populations except in one sample. In this population from 3 samples analysed, two samples contained 3 subunits, but one of sample showed an extra band at top (1Dy) (Fig. 1).
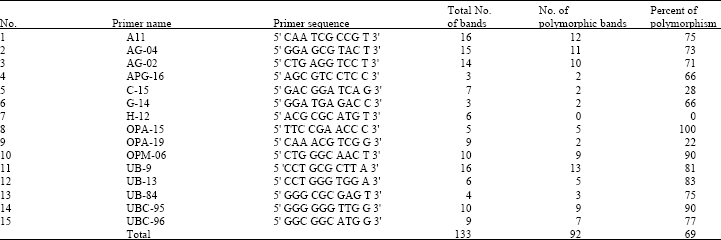
RAPD analysis: In the RAPD analysis, 15 ten-mer primers were used to amplify all of the genotypes; 14 of these showed informative polymorphic products resolvable by gel electrophoresis. A total of 133 bands were screened (average of 8.9 bands per primer) among which 92 were polymorphic (69%). The number of fragments generated per primer varied between 3 and 16. The highest and the lowest number of polymorphic bands per assay unit were 0 (primer OPA-15) and 13 (primer UB-9), respectively (Table 2). Figure 2 shows an example of such a typical RAPD pattern using the WMS-43 primer.
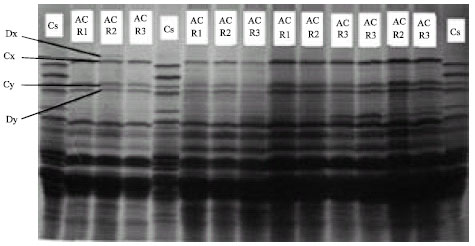 | |
| Fig. 1: | Example of High Molecular Weight (HMW) subunits of glutenin among some populations of Ae. cylindrica. Cs is chines spring; R1, R2 and R3 are three samples taken from each population |
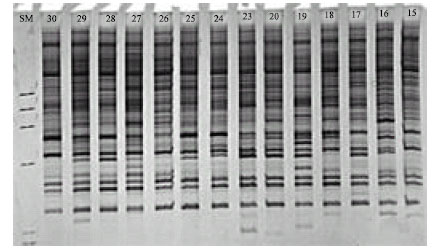 | |
| Fig. 2: | Example of polymorphism between some populations of Ae. cylindrica as revealed by RAPD primer WMS-43 |
| Table 2: | RAPD primers with sequence, total number of bands, number of polymorphic bands, percent of polymorphism |
 | |
Estimates of genetic similarity of RAPD based on 92 polymorphic markers between 28 populations of Ae. cylindrica ranged from 0.41 for AC17/AC24, AC13/AC17 and AC12/AC15 to 0.84 for AC20/AC23 with an average of 0.63.
Genetic similarity values were used for cluster analysis through UPGMA, resulting in a dendrogram (Fig. 3). Cluster analysis revealed that populations Ac7 and Ac17 from Mahabad and Shahrod origion sites, respectively were more genetically distinct from the others (Fig. 1) and the most obvious pairs were Ac20 and Ac23. PCO analyses revealed that for RAPD data the first two components of the PCO explained 14.8 and 9.1% of the total variation (Fig. 4).
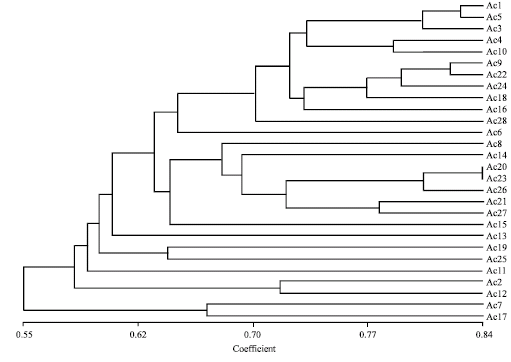 | |
| Fig. 3: | UPGMA dendrogram showing genetic relationships among the 28 populations of Ae. cylindrica used in this study. The dendrogram was constructed based on genetic similarity calculated according to Dice coefficient. For population abbreviations see Table 1 |
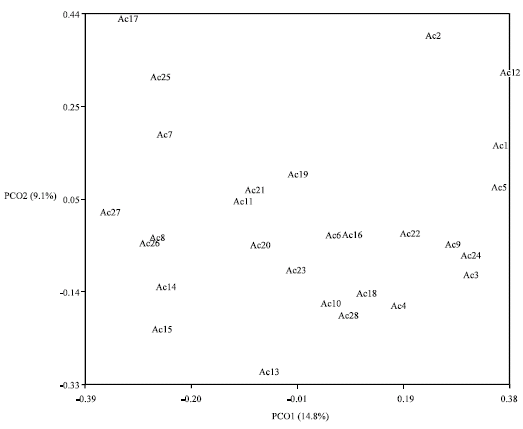 | |
| Fig. 4: | Patterns of relationships among 28 populations of Ae. cylindrica revealed by principal coordinate analysis based on RAPD data. For population abbreviations see Table 1 |
Although the results of PCO didn’t correspond totally to those from cluster analysis, but it confirmed some subgrouping obtained by cluster analysis.
DISCUSSION
In this study, two molecular methods, seed storage protein and RAPD, were used to investigate the genetic diversity in the 28 populations of Ae. cylindrica. Seed storage protein analysis showed very low genetic diversity in studied populations. Isozyme analysis also conducted by Watanabe et al. (1994) and Hegde et al. (2002) revealed little or no variation among accessions of jointed goatgrass. We could find an extra band at top (1Dy) that it was very similar to top band of Ae. triuncialis. As this species distributes frequently with Ae. triuncialis along roadsides, therefore it is possible that the accession with extra band is a hybrid between Ae. triuncialis and Ae. cylindrica.
Compare to seed storage protein, RAPD markers revealed more polymorphic fragments. This is expected, as proteins markers reflect only variation in the coding parts of the genome, which is by nature more conservative and thus less polymorphic. While RAPDs can detect variation in both coding and non coding sequences, and the length of the primers allows the amplification of a large number of fragments with a single primer (Guadagnuolo et al., 2001). The RAPD technique has been widely applied in studies of wheat genetic diversity (Cao et al., 1998; Sun et al., 2003; Naghavi et al., 2004; Ahmad Khan et al., 2005) and has shown its usefulness in rapid detection of genetic variation. Although the level of RAPD polymorphism within wheat itself has been reported to be low (Devos and Gale, 1992), but the level of RAPD polymorphism in wild wheat has been found to be higher than that in common bread wheat (Sun et al., 2003).
RAPD amplifications provided the largest set of polymorphic markers. In this study, 92 RAPD loci were used to investigate the genetic diversity in the populations of Ae. cylindrica. The amount of polymorphism found in this research (69%) was more than what reported in previous studies (Okuno et al., 1998, Guadagnuolo et al., 2001; Pester et al., 2003). These differences might be related to the utilization of different Ae. cylindrica germplasms as well as the used of different primers sequences.
In this research we found that there was little relationship between genetic divergence and geographical origins, so that the populations from similar geographical places (AC1 with AC11 or AC27) belonged to separate clusters. Conversely, populations from different geographical conditions (such as AC7 and AC17) relatively tended to be clustered in one part of the dendrogram.
In conclusion, our results show that compare to seed storage protein, RAPD is suitable for genetic diversity assessment in Ae. cylindrica populations and it appears to be the most suitable for this kind of study. This technique has the advantage of requiring very small quantities of template DNA and no prior knowledge of sequencing in the target genome (Pester et al., 2003). Even if the dominance of RAPD markers remains a problem for population genetic studies, it is possible to overcome it, if specific fragments for each taxon or group of taxa compared are produced (Guadagnuolo et al., 2001).
REFERENCES
- Ahmad Khan, I., F.S. Awan, A. Ahmad, Y. Fu and A. Iqbal, 2005. Genetic diversity of Pakistan wheat germplasm as revealed by RAPD markers. Genet. Resour. Crop Evol., 52: 239-244.
Direct Link - Cao, W., P. Hucl, G. Scoles and R.N. Chibbar, 1998. Genetic diversity within spelta and macha wheats based on RAPD analysis. Euphytica, 104: 181-189.
CrossRefDirect Link - Galaev, A.V. and Y.M. Sivolap, 2005. Molecular-genetic analysis of wheat (T. aestivum L.) genome with introgression of Ae. Cylindrica host genetic elements. Cytol. Genet., 39: 57-66.
Direct Link - Gill, B.S., B. Friebe, W.J. Raupp, D.L. Wilson and T.S. Cox et al., 2006. Wheat genetics resource center: The first 25 years. Adv. Agro., 89: 73-136.
Direct Link - Hegde, S.G., J. Valkoun and J.G. Waines, 2002. Genetic diversity in wild and weedy Aegilops, Amblyopyrum and Secale species-a preliminary survey. Crop Sci., 42: 608-614.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Naghavi, M.R., M. Mardi, H.A. Ramshini and B. Fazelinasab, 2004. Comparative analyses of the genetic diversity among bread wheat genotypes based on RAPD and SSR markers. Iranian J. Biotechnol., 2: 195-202.
Direct Link - Nei, M. and W.H. Li, 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA., 76: 5269-5273.
CrossRefPubMedDirect Link - Saghai-Maroof, M.A., K.M. Soliman, R.A. Jorgensen and R.W. Allard, 1984. Ribosomal DNA spacer-length polymorphisms in barly: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci., 81: 8014-8018.
CrossRefDirect Link - Sun, G., M. Bond, H. Nass, R. Martin and Z. Dong, 2003. RAPD polymorphisms in spring wheat cultivars and lines with different level of Fusarium resistance. Theor. Applied Genet., 106: 1059-1067.
CrossRefDirect Link