
F. Najafi
Department of Biology, Tarbiat Moallem University, P.O. Box 15815-3587, Tehran, Iran
R. A. Khavari-Nejad
Department of Biology, Tarbiat Moallem University, P.O. Box 15815-3587, Tehran, Iran
F. Rastgar-jazii
National Research Center for Genetic Engineering and Biotechnology,
P.O. Box 14155-6343, Tehran, Iran
M. Sticklen
Department of Crop and Soil Science, Michigan State University, Plant and Soil Science Building, East Lansing, Michigan 48 824, USA
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 16 | Page No.: 2752-2755







ABSTRACT
The effects of salt stress were studied on growth and physiology of pea (Pisum sativum L. cv. Green Arrow) in a pot study. Pea plants were treated with NaCl at 0, 10, 30, 50 and 70 mM in Hoagland solution. Plants were harvested after 21 days for measurements of physiological parameters. The highest NAR and RGR were found in 10 mM NaCl. However, in 70 mM NaCl, RGR and RLGR were significantly decreased in respect of other concentrations of NaCl. In 50 and 70 mM NaCl, chlorophylls contents and photosynthetic rate, were significantly decreased and CO2 compensation concentration and respiration rate increased in comparison with control. In 10 and 30 mM NaCl gas exchanges and chlorophyll contents were not significantly decrease in respect of control. Results indicated that Pisum sativum L. cv. Green Arrow can tolerate to 70 mM NaCl, also growth of plants in 10 and 30 mM NaCl was better than that of those in 0 mM NaCl.
PDF Abstract XML References Citation
How to cite this article
F. Najafi, R. A. Khavari-Nejad, F. Rastgar-jazii and M. Sticklen, 2007. Growth and Some Physiological Attributes of Pea (Pisum sativum L.) As Affected by Salinity. Pakistan Journal of Biological Sciences, 10: 2752-2755.
DOI: 10.3923/pjbs.2007.2752.2755
URL: https://scialert.net/abstract/?doi=pjbs.2007.2752.2755
DOI: 10.3923/pjbs.2007.2752.2755
URL: https://scialert.net/abstract/?doi=pjbs.2007.2752.2755
INTRODUCTION
Plants are influenced by a variety of biotic or abiotic stresses, such as drought, salt loading, and freezing as changes in their development, growth and productivity. One of the major abiotic stresses that affect plant productivity is water stress resulting from drought or salinity (Gueta-Dahan et al., 1997).
There is increasing evidence that salinity changes photosynthetic parameters, including osmotic and leaf water potential, transpiration rate, leaf temperature and relative leaf water content. Salinity can seriously change the photosynthetic carbon metabolism, leaf-chlorophyll content, as well as photosynthetic efficiency (Seemann and Critchley, 1985; Sharkey, 1985). Changes in these parameters depend on the severity and duration of stress (Lakshmi et al., 1996; Misra et al., 1997) and on plant species (Dubey, 1994).
The effect of salinity on growth, nodulation and N2 fixation of legumes plants has been studied (Lauter and Munns, 1986; Elsheikh and Wood, 1990). The adaptation to salinity stress is accompanied by alternations in the levels of numerous metabolites, proteins and mRNA (Serrano, 1996). Various genes, expression of which is activated in response to salt stress, have been identified (Shinozaki, 1999; Kawasaki et al., 2001). Some of these genes encode for protective proteins such as osmotin (Zhu et al., 1995), late embryogenesis abundant (LEA) proteins (Espelund et al., 1992) and ion transporters (Blumwald, 2000) others code for enzymes that participate in metabolic processes specifically triggered by salinity stress (Cushman, 1992; Gong et al., 2001).
In this research, physiological changes of pea plants in saline environment are studied. Results indicated that, in 10 and 30 mM NaCl, pea plants have more growth rate than control (0 mM NaCl) plants and also they can tolerant to 70 mM NaCl.
MATERIALS AND METHODS
Pea (Pisum sativum L. cv. Green Arrow) seeds were prepared from Agricultural Research Center, IRAN. Pea seeds were surface-sterilized in 70% (v/v) ethanol (1 min) followed by 1% (w/v) sodium hypochorite (20 min) and washed five times with sterile distilled water (Schroeder et al., 1993).
Seeds germinated in pots containing perlite in a growth chamber under a 24°C temperature and at a relative humidity of 70%. Germinated seeds were translated to pots in growth chamber with 17 h light periods and 300 μmol quanta m-2 sec-1 light intensity, day/night temperatures of 25/18°C and irrigated with a Hoagland´s solution. After 10 days, the seedlings were transplanted in the saline nutrient solutions containing 0, 10, 30, 50 and 70 mM sodium chlororide, with pH 6.5 and fresh nutrient solution replaced the old every week. The plants were grown under controlled environment (17 h light periods, 300 μmol quanta m-2 sec-1 light intensity, day/night temperatures of 25/18°C) in a greenhouse. After 21 days of experimental duration, for per physiological analysis from per treatment four plants were harvested.
Chlorophylls a and b and total chlorophylls were estimated as described by Arnon (1949). Photosynthetic rate (Pn), respiration rate and CO2 compensation concentration ( Γ ) were determined from intact plants, employing an Infrared Gas (CO2) Analyzer (IRGA) as described by Khavari Nejad (1980, 1986). Growth analyses were carried out using the equations of Watson (1952) and Evans and Hughes (1962). The research was conducted using completely randomized design with four replications. Data were analyzed statistically using SAS software.
RESULTS
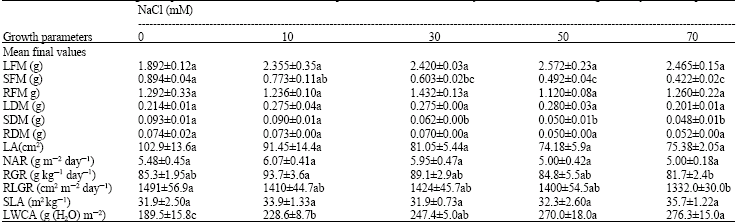
Growth: Fresh and dry weight of the stem in 50 and 70 mM NaCl were significantly lower than those of plants treated with 0, 10 and 30 mM NaCl. Increasing fresh and dry weight of leaf in 10 and 30 mM NaCl were not significant in respect of control.
NAR, RLGR and SLA were not significantly decreased in different treatments of NaCl. However, LWCA in 50 and 70 mM NaCl was higher than that of plants in 0, 10 and 30 mM NaCl also RGR was significantly decreased in 70 mM NaCl (Table 1).
Gas exchange: In 70 mM NaCl, Pn was significantly decreased in respect of other treatments. However, Pn was not significantly decreased in 10, 30 and 50 mM NaCl in respect of control. Respiration rate in 50 and 70 mM NaCl was higher than that of plants in 0, 10 and 30 mM NaCl. CO2 compensation concentration with increasing concentration of NaCl was not significantly enhanced (Table 2 ).
Chlorophylls contents: Concentration of chlorophylls of leaves in 50 and 70 mM NaCl were significantly decreased in respect of 0, 10 and 30 mM NaCl. In 70 mM NaCl Chl.a/ Chl.b ratio was decreased in respect of other treatments (Table 3).
DISCUSSION
Decreasing effects of NaCl on growth have been reported in legumes, as well as the chick-pea (Dua, 1992), soybean (Singleton and Bohlool, 1984) and faba bean (Zahran and Sprent, 1986; Cordovilla et al., 1996). In the present work, the effect of NaCl on pea plants were investigated and indicated that Pisum sativum L. cv. Green Arrow was tolerant to 70 mM NaCl and growth in 10 and 30 mM were increased in respect of control.
Gas exchanges were more sensitive to salinity than growth parameters. Amongst damages caused by salinity stress in plants, the reduction of photosynthetic processes is one of the most important (Delfine et al., 1999). Our results indicated that in 70 mM NaCl, Pn, was significantly decreased and respiration rate and Γ were increased in respect of control. However, 10, 30 and 50 mM NaCl had a low effect on gas exchanges (Table 2).
| Table 1: | Effects of NaCl on growth parameters, Means (±SE) of four replicates, Numbers followed by the same letter are not significantly different (p>0.05) |
 | |
| Table 2: | Effects of NaCl on photosynthetic rate (Pn), respiration rate, CO2 compensation concentration (Γ), Means (±SE) of four replicates, Numbers followed by the same letter are not significantly different (p>0.05) |
 | |
| Table 3: | Effects of NaCl on chlorophylls content and chl.a/chl.b ratio, Means (±SE) of four replicates, Numbers followed by the same letter are not significantly different (p>0.05) |
 | |
Growth parameters are ultimately a function of total photosynthetic capacity and comparison of photosynthetic and growth responses to salinity illustrate that with decreasing photosynthetic rate in 70 mM NaCl, RGR and RLGR are significantly decreased in respect of another concentrations of NaCl.
LWCA was significantly enhanced in all treatments. As described earlier (Tester and Davenport, 2003) an ability to grow in saline conditions has been attributed to an ability to close stomata. In fact both glycophytes and halophytes tend to show reduced stomatal conductance in high NaCl conditions (Ball, 1988; Robinson et al., 1997; James et al., 2002). The productivity of crops in saline soils is limited to a large extent by the low rate of gas exchange imposed by the drought component of NaCl stress and further reduction in stomatal conductance and photosynthetic rate is unlikely to improve plant productivity (Tester and Davenport, 2003).
In this study, in plants treated with 50 and 70 mM NaCl, chlorophylls contents were significantly decreased as compared with control. However, decreasing chlorophylls contents in plants grown in 10 and 30 mM NaCl were not significant, the decline in chlorophyll a/b ratio of plants treated with 70 mM NaCl was significantly higher than that of plants treated in others treatments. Decrease in chlorophyll concentration in salinized plants could be attributed to increased activity of chlorophyll-degrading enzyme chlorophyllase (Reddy and Vora, 1986).
It is concluded that pea plants ( Pisum sativum L. cv. Green Arrow) are resistant to 50 and 70 mM NaCl and their growth is higher in 10 and 30 mM NaCl than that of plants grown in 0 mM NaCl.
REFERENCES
- Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol., 24: 1-15.
CrossRefPubMedDirect Link - Ball, M.C., 1988. Salinity tolerance in the mangroves Aegiceras corniculatum and Avicennia marina. I. Water use in relation to growth, carbon partitioning and salt balance. Aust. J. Plant Physiol., 15: 447-464.
CrossRefDirect Link - Blumwald, E., 2000. Sodium transport and salt tolerance in plants. Curr. Opin. Cell Biol., 12: 431-434.
CrossRefDirect Link - Cordovilla, M.P., F. Ligero and C. Lluch, 1996. Growth and nitrogen assimilation in nodules in response to nitrate levels in Vicia faba under salt stress. J. Exp. Bot., 47: 203-210.
Direct Link - Espelund, M., S. Saeboe-Larssen, D.W. Hughes, G.A. Galau, F. Larsen and K.S. Jakobsen, 1992. Late embryogenesis-abundant genes encoding proteins with different numbers of hydrophilic repeats are regulated differentially by abscisic acid and osmotic stress. Plant J., 2: 241-252.
CrossRefDirect Link - Evans, G.C. and A.P. Hughes, 1962. Plant growth and the aerial environment. III. On the computation of unit leaf rate. New Phytol., 61: 322-327.
Direct Link - Gong, Z., H. Koiwa, M.A. Cushman, A. Ray and D. Bufford et al., 2001. Genes that are uniquely stress regulated in Salt Overly Sensitive (SOS) mutants. Plant Physiol., 126: 363-375.
Direct Link - Gueta-Dahan, Y., Z. Yaniv, B.A. Zilinskas and G. Ben-Hayyim, 1997. Salt and oxidative stress: Similar and specific responses and their relation to salt tolerance in citrus. Planta, 204: 460-469.
CrossRef - James, R.A., A.R. Rivelli, R. Munns and S. von Caemmerer, 2002. Factors affecting CO2 assimilation, leaf injury and growth in salt-stressed durum wheat. Functional Plant Biol., 29: 1393-1403.
Direct Link - Kawasaki, S., C. Borchert, M. Deyholos, H. Wang and S. Brazille et al., 2001. Gene expression profiles during the initial phase of salt stress in rice. Plant Cell., 13: 889-906.
PubMedDirect Link - Lakshmi, A., S. Ramanjulu, K. Veeranjaneyulu and C. Sudhakar, 1996. Effect of NaCl on photosynthesis parameters in two cultivars of mulberry. Photosynthetica, 32: 285-289.
Direct Link - Misra, A.N., S.M. Sahu, M. Misra, P. Singh and I. Meera et al., 1997. Sodium chloride induced changes in leaf growth and pigment and protein contents in two rice cultivars. Biol. Plant., 39: 257-262.
CrossRefDirect Link - Reddy, M.P. and A.B. Vora, 1986. Changes in pigment composition, Hill reaction activity and saccharides metabolism in Bajra (Pennisetum typhoides S & H) leaves under NaCl salinity. Photosynthetica, 20: 50-55.
Direct Link - Robinson, M.F., A.A. Very, D. Sanders and T.A. Mansfield, 1997. How can stomata contribute to salt tolerance? Ann. Bot., 80: 387-393.
CrossRefDirect Link - Schroeder, H.E., A.H. Schotz, T. Wardley-Richardson, D. Spencer and T.J.V. Higgins, 1993. Transformation and regeneration of two cultivars of pea (Pisum sativum L.). Plant Physiol., 101: 751-757.
Direct Link - Seemann, J.R. and C.H. Critchley, 1985. Effects of salt stress on the growth, ion content, stomatal behaviour and photosynthesis capacity of a salt-sensitive species, Phaseolus vulgaris L. Planta, 164: 151-162.
CrossRef - Serrano, R., 1996. Salt tolerance in plants and microorganisms: Toxicity targets and defense responses. Int. Rev. Cytol., 165: 1-52.
CrossRefDirect Link - Sharkey, T.D., 1985. Photosynthesis in intact leaves of C3 plants: Physics, Physiology and rate limitations. Botanic. Review, 51: 53-105.
CrossRef - Watson, D.J., 1952. The physiological basis of variation in yield. Adv. Agron., 4: 101-145.
CrossRefDirect Link - Zhu, B., T.H. Chen and P.H. Li, 1995. Expression of three osmotin-like protein genes in response to osmotic stress and fungal infection in potato. Plant Mol. Biol., 28: 17-26.
CrossRefDirect Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link