
Mohamad K. Nusier
Department of Biochemistry and Molecular Biology,
Hameed N. Bataineh
Department of Physiology, Jordan University of Science and Technology, School of Medicine, Irbid 22110, Jordan
Haytham M. Daradka
Department of Biology, Faculty of Science, Jarash University, Jarash 21122, Jordan
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 16 | Page No.: 2728-2731







ABSTRACT
Ingestion of propranolol at 10 and 15 mg kg-1 body weight for 35 days by adult male mice was investigated for its effects on fertility. Body weight and absolute and relative testes weights were reduced and the average weights of epididymis, ventral prostate and seminal vesicle decreased significantly. A significant decline of spermatogenesis in testes due to a decrease in the number of primary, secondary spermatocytes and spermatids in the treatment group 2 (15 mg kg-1) is attributed to a significant decrease in testosterone, LH and FSH. Sperm motility and density were also significantly decreased in the cauda epididymis and in the testes of group 2 treated male mice. In addition, the treatment markedly increased the number of fetal resorptions in female mice impregnated by the group 2 males, thereby reducing their fertility.
PDF Abstract XML References Citation
How to cite this article
Mohamad K. Nusier, Hameed N. Bataineh and Haytham M. Daradka, 2007. Adverse Effects of Propranolol on Reproductive Function in Adult Male Mice. Pakistan Journal of Biological Sciences, 10: 2728-2731.
DOI: 10.3923/pjbs.2007.2728.2731
URL: https://scialert.net/abstract/?doi=pjbs.2007.2728.2731
DOI: 10.3923/pjbs.2007.2728.2731
URL: https://scialert.net/abstract/?doi=pjbs.2007.2728.2731
INTRODUCTION
β-adrenoceptor antagonist are widely used in cardiovascular medicine (Bhatt et al., 2007). They protect the heart by blocking the effect of adrenaline to maintain a normal heart beat and normal blood pressure (Ojeda et al., 2002; Zelaszczyk and Kiec-Kononowicz, 2007). Propranolol has been associated with male and female sexual side effects (Adamowicz and Kala, 2005; Ko et al., 2002). It has been previously reported that systemic administration of β-adrenoceptor antagonists differentially alters sexual function in male rats (Smith et al., 1990). Specifically, the nonselective β-adrenoceptor antagonists inhibited male rat sexual behavior (Smith et al., 1990), while selective β1-drenoceptor antagonists did not (Smith et al., 1990; Weiner, 1985). The sexual inhibition was evidenced by the decrease in the number of males copulating to ejaculation and by increases in the intervals between copulatory events and a decreased copulatory efficacy. Propranolol effects were not only limited to copulation, but also to deficits in erectile and ejaculatory reflexes (Smith et al., 1995).
Therefore, the present research was designed to investigate the detailed effects of propranolol on sexual maturation, fertility and testicular functions in male Albino mice.
MATERIALS AND METHODS
Thirty adult male and 60 female Albino mice weighing approximately 40 g were bred in the Animal House Unit at JUST, School of Medicine, between January and February 2007. Mice were maintained at a controlled temperature of 21±1°C and under a 12-h-light: 12-h-dark schedule. Food and water were supplied ad libitum.
Generic propranolol was purchased from a local pharmacy. This compound was then dissolved in distilled water and administered orally to mice using animal feeding intubation needles (Popper and Sons, New York) in a concentration of 10 and 15 mg kg-1 body weight (1 mL volume) as single daily doses. Similarly, the controls received gastric infusions of 1 ml distilled water, the same way the experimental mice did.
Male mice were randomly assigned to control or experimental groups. Experimental male mice were divided into 2 groups: group 1 and 2 were fed 10 and 15 mg kg-1 body weight propranolol for 35 days, respectively. All male mice were healthy and continued to receive their respective drinking water and food throughout the experimental period.
To estimate the fertility in both experimental and control male mice; each male was placed in an individual cage with two virgin untreated females of the same strain for 10 days, brought into estrus by sequential subcutaneous treatment with 0.9 mg of estradiol benzoate (Sigma Chemical Co. St Louis, MO, USA) 54 h before testing and 0.1 mg of progesterone (Gift from Roussel Uclaf, Paris, France) 6 h before testing. The hormones were dissolved in corn oil (ALFCO: Arab International Food and Oil Processing Co.) in a total volume of 0.1 mL.
They were left together for 10 days during which two estrus cycles should have elapsed (Rugh, 1968). One week after the removal of the males, the females were killed by cervical dislocation under light ether anesthesia and the number of pregnant females, implantation sites, viable fetuses and fetal resorptions were recorded (Prasad et al., 1972).
All males were sacrificed after 35 days of exposure. Blood was collected by heart puncture using a sterile syringe and serum was separated and stored at -20°C for biochemical analysis. Body weight and weights of paired testes and seminal vesicles (stripped of fluid) were recorded. The reproductive organs of male mice including the testes, epididymides, ventral prostrate, seminal vesicle and vas deferens were fixed in Bouin’s fixative for histological studies.
The sperm motility and sperm counts of cauda epididymis were determined by the method described earlier Quantitative motility (%) was determined by counting both motile and immotile spermatozoa per unit area. Cauda epididymis and testicular sperm counts were performed by routine procedure and expressed as millions mL-1 of suspension (Abercrombie, 1946). Histological evaluation and sperm analyses were performed by a person who is unfamiliar with mice groups in this experiment. The Bouin's fixed reproductive organs were processed for paraffin embedding, sectioning (5 μm) and staining (Harris haematoxylin and eosin).
Using Camera Lucida, 100 circular appearing seminiferous tubules were traced at x80 and the diameter of each tubule was measured separately. The measurement was expressed in terms of mean of all the traced tubules. Similarly, Leydig cell nuclei were traced at x800. The epithelial cell height of caput and cauda epididymis and seminal vesicle were also traced at x 360.
The spermatogonia, spermatocytes and spermatids were counted in 5 μm thick cross sections of 10 seminiferous tubules in 10 animals of each group. All raw counts were transformed into true counts by an adaptation of Abercrombie formula {T = C (s/s+dx) 100 [T = True counts; C = Crude counts; S = Thickness of section; d = Diameter of nuclei]} (Dixon, 1975) from germ cell diameter measurement. Interstitial cell types (such as fibroblast, immature and mature Leydig cells and degenerating cells) were counted, applying a differential count over a 200-cell population and statistically verified by the binomial distribution (Ipstein and Poly, 1970).
Serum testosterone concentration was measured by Enzyme Immunoassay Kits (Cayman Chemical, USA). Serum LH (Luteinizing Hormone) and FSH (Follicle Stimulating Hormone) concentrations were measured by a mice LH enzyme immunoassay and mice FSH enzyme immunoassay systems (Amersham, England), respectively.
Data were expressed as mean±SD and Median (statistical package for social sciences [SPSS, version 11.5]. Differences between control and propranolol exposed male groups were analyzed using either the Chi-square test or Student t-test. A p-value of <0.05 was considered statistically significant (Ipstein and Poly, 1970).
RESULTS
Table 1 shows that intragastric administration of proppranolol significantly decreased body weight of treated males when compared to those of control group. Furthermore, their relative weights of testes, epididymides, seminal vesicles, ventral prostate and vas deferens were significantly reduced in group 2.
Table 2 shows that sperm motility in cauda epididymis, sperm density and serum levels of testosterone, FSH and LH were significantly decreased in group 2 treated animals in comparison with controls.
Table 3 shows that administration of propranolol caused a significant decrease in the germinal cell population: spermatocytes (primary and secondary) and spermatids were decreased to a significantly lower level in group 2. Furthermore, in group 1 there was a significant decrease in secondary sprematocytes, spermatids, fibroblasts, immature Leydig cells and mature Leydig cells. Similarly, the numbers of fibroblasts, immature and mature
| Table 1: | Body and organ weights of male mice ingested propranolol |
 | |
| Results are expressed as mean±SD. *p<0.05, **p<0.01 significantly different from control group (Student’s t-test). Group 1 received propranolol 10 mg kg-1 body weight. Group 2 received propranolol 15 mg kg-1 body weight | |
| Table 2: | Fertility parameters and sperm dynamics of male mice ingested propranolol |
 | |
| Results are expressed as mean±SD.*p<0.05, **p<0.001 significantly different from control group (Student’s t-test). Group 1 received propranolol 10 mg kg-1 body weight. Group 2 received propranolol 15 mg kg-1 body weight | |
| Table 3: | Testicular cell population dynamics of male mice ingested propranolol |
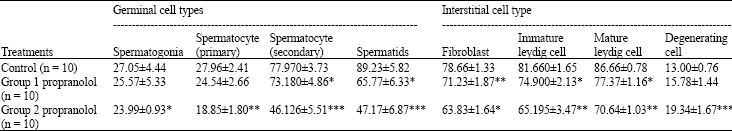 | |
| Results are expressed as mean±SD. *p<0.05, **p<0.001, ***p<0.001 significantly different from control group (Student’s t-test). Group 1 received propranolol 10 mg kg-1 body weight. Group 2 received propranolol 15 mg kg-1 body weight | |
| Table 4: | Effect of propranolol ingestion on fertility in adult male mice |
 | |
| Results are expressed as mean±SD. *p<0.05, **p<0.001, significantly different from control group (Student’s t-test and Chi-square test ξ). Group 1 received propranolol 10 mg kg-1 body weight. Group 2 received propranolol 15 mg kg-1 body weight | |
Leydig cells were also significantly decreased in group 2. The number of degenerating cells, however, was significantly increased in group 2.
Table 4 shows that propranolol exposure reduced fertility as indicated by the smaller number of pregnant females impregnated by the propranolol-exposed males. However, the number of implantations and number of viable fetuses were significantly decreased in those females impregnated by groups 1 and 2 males. The total number of resorptions was also significantly increased in females impregnated by group 2 males.
DISCUSSION
The animal model used in this work has previously been used to assess the effect of various extracts obtained from medicinal plants on reproductive functions in male (Nusier and Bataineh, 2005; Bataineh and Nusier, 2005; Bataineh et al., 1998).
Spermatogenic process in mice requires 35 days, out of which spermatozoa spend the last 6 to 7 days in the final transit through epididymides (Ke and Tso, 1982). Propranolol was administrated for one complete spermatogenic cycle.
The present investigation shows that oral administration of propranolol reduced fertility in male albino mice. The weights of reproductive organs were markedly decreased. The weight, size and secretory function of testes, epididymis, seminal vesicles, ventral prostate and vasa deferentia are closely regulated by androgens (Choudhary and Steinberger, 1975). The process of spermatogenesis and accessory reproductive organs function are androgen dependent (Agrawal et al., 1986; Dym et al., 1979). Histological studies of testes, epididymis, seminal vesicles, ventral prostate and vasa deferentia also confirmed the results (Agrawal et al., 1986). The drug may act on pituitary gland and decrease the main hormone of spermatogenesis (Agrawal et al., 1986). Decrease androgen production is reflected by decreased number of mature Leydig cells and their functional status. In the present study, the number of degenerating Leydig cells was significantly increased; this may reflect the decrease of androgen levels. It was further confirmed by the decreased number of spermatocytes (primary and secondary) and spermatids as these stages are completely androgen dependent (Agrawal et al., 1986). The decreased weight and histometry of reproductive organs further confirmed the decrease in androgen levels. A significant increase in the sperm motility of cauda epididymis was observed in the treated group.
Results presented in this manuscript also showed that the ingestion of propranolol by adult male mice caused a slight decrease in the number of females impregnated by the treated males. However, the number of implantations sites and the number of viable fetuses were reduced. These observations may be due to the decrease in sperm motility and function. Reduction in implantation and phytotoxic effects might be due to cytotoxic effects which can result in decreased fertility, failure of preimplantation or postimplantation death. Cytotoxic agents can disrupt pregnancy possibly by interfering with mitotic division of the fetus (Desjardins, 1978; Working et al., 1985).
The increased resorption is further evidence, along with the reduced implantation rate, that propranolol at 15 g kg-1 day-1 is having an adverse effect on male sperm.
These results may suggest that propranolol imposes toxic effects on fertility in male mice.
ACKNOWLEDGMENT
This research project was supported by the Deanship of Research at Jordan University of Science and Technology, School of Medicine.
REFERENCES
- Abercrombie, M., 1946. Estimation of nuclear population from microtome section. Anatomical Rec., 94: 239-247.
CrossRefDirect Link - Agrawal, S., S. Chauhan and R. Mathur, 1986. Antifertility effects of embelin in male rats. Andrologia, 18: 125-131.
CrossRefPubMedDirect Link - Bataineh, H., M.H. Al-Hamood and A.M. Elbetieha, 1998. Assessment of aggression, sexual behavior and fertility in adult male rat following long-term ingestion of four industrial metals salts. Hum. Exp. Toxicol., 17: 570-576.
CrossRefDirect Link - Bataineh, H. and M. Nusier, 2005. Effect of cholesterol diet on reproductive functions in male albino rats. Saudi Med. J., 26: 398-404.
Direct Link - Bhatt, L.K., K. Nandakumar, S.L. Bodhankar, J. Bansal and P. Piplani, 2007. Beta-blocking activity of PP-34, a newly synthesized aryloxypropanolamine derivative and its cardioprotective effect against ischemia/reperfusion injury in laboratory animals. J. Pharm. Pharmacol., 59: 429-436.
PubMedDirect Link - Ke, Y.B. and W.W. Tso, 1982. Variations of gossypol susceptibility in rat spermatozoa during spermatogenesis. Int. J. Fertil., 27: 42-46.
PubMedDirect Link - Ko, D.T., P.R. Hebert, C.S. Coffey, A. Sedrakyan, J.P. Curtis and H.M. Krumholz, 2002. β-blocker therapy and symptoms of depression, fatigue and sexual dysfunction. J. Am. Med. Assoc., 288: 351-357.
CrossRefDirect Link - Ojeda, R.R., J.C. Sanchez, A.O. Fernandez, J.M.D. Roldan and A.H. Fernandez, 2002. Propranolol protects the myocardium and prevents arterial hypotension in an experimental organ donation model. Rev. Esp. Anestesiol. Reanim., 49: 229-237.
PubMedDirect Link - Zelaszczyk, D. and K. Kiec-Kononowicz, 2007. Biocatalytic approaches to optically active beta-blockers. Curr. Med. Chem., 14: 53-65.
PubMedDirect Link