
Zohreh Hosseinzadeh
Department of Biology,
Ahmad Ali Moazedi
Department of Biology,
Rahim Chinipardaz
Department of Statistics and Mathematics, Shahid Chamran University, Ahwaz, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 16 | Page No.: 2653-2658







ABSTRACT
The aim of the present study is for evaluating the effect of different doses of palmitic acid on spatial learning in T-maze. So, Wister adult male rats were used in five groups and considered 10 rats for each group. The first group as a control group was fed with ordinary diet. Four groups were fed with a diet containing palmitic acid (10%) for 1, 2, 3 and 4 weeks. then rats were trained for spatial learning task by using T-maze at subsequently 9 days based on standard method. The result showed that, spatial learning increased by diet containing palmitic acid 10% for 1, 2 and 3 weeks were significant (p<0.05). However, dietary palmitic acid effect on learning because palmitic acid via palmitoylation modifies numerous important neuronal protein including GAP-43, NMDA receptor, AMPA receptor and these receptors are critical role in learning.
PDF Abstract XML References Citation
How to cite this article
Zohreh Hosseinzadeh, Ahmad Ali Moazedi and Rahim Chinipardaz, 2007. The Effect of Palmitic Acid on Spatial Learning and Extinction in Adult Male Rat. Pakistan Journal of Biological Sciences, 10: 2653-2658.
DOI: 10.3923/pjbs.2007.2653.2658
URL: https://scialert.net/abstract/?doi=pjbs.2007.2653.2658
DOI: 10.3923/pjbs.2007.2653.2658
URL: https://scialert.net/abstract/?doi=pjbs.2007.2653.2658
INTRODUCTION
The fatty acid composition of brain was measured to determine the relationship between the resulting range in brain fatty acid composition and performance on the working memory task in morris water maze (Wainwright et al., 1998). The relation between a biological factor (fatty acids, FA) and a cognitive processing speed factor (temporal processing acuity, TPA) that are both suggested to relate to neuronal and cognitive functioning (Laasonen et al., 2006).
n-3polyunsaturated fatty acids (linoleate and linolenate) and saturated fatty acids (palmitate, stearate) and monounsaturated fatty acids (oleate) were supplied in the diet to artificially reared rat pups over the period of the brain growth spurt and the onset of myelinogenesis (Edmond et al., 1998).
The promising relationships between epilepsy and seizure on one hand and fatty acids on the other hand complex and not obvious (Mostosky et al., 2004).
Few topic in nutrition cause as much controversy and concern and are as frequently misunderstood, as fat (Taubes, 2001). The reported cautionary warning from the medical profession has been to dramatically reduce the amount of fat one eats, so as to minimize the risks associated with cardiovascular diseases, diabetes and other chronic disorders (Kodas et al., 2004). Fatty acids deficiency may impair normal neurological development (Yehuda et al., 2006).
However deficiencies in fat intake are equally likely to contribute to health hazards, including increased risk of infection, dysregulation of cyclic and rhythmic activity and impairment cognitive and sensory function (especially infants) (Ruthrich et al., 1999).
A consensus has emerged from recent research that it is not so much amount of fat we eat as it is the balance of the different type of fats (Yehuda, 2003, 2002).
The type of dietary fat we consume affects the biology of each cell and determines how well it can perform its vital function and its ability to resist diseases (Ruthrich et al., 1999). Indeed among the significant component of cell membranes are the phospholipids, which contain fatty acids. The type of fatty acids in the diet determines the type of fatty acids that is available to the composition of cell membranes. A phospholipids made from a saturated fat has a different structure and is less fluid, than incorporates an unsaturated and essential fatty acid (Yehuda, 2003; McCann and Ames, 2007). Based on different reports about the effects of saturated fatty acids spatially palmitic acid on nerve system and learning and memory and in past studies our laboratory showed that compare between groups fed dietary contain butter (10%) for 1, 2 and 3 weeks indicated enhanced learning performance that may be cause presence of saturated fatty acids in butter and also the most amount of saturated fatty acids in the butter is palmitic acid. So, in this study the effects of palmitic acid on spatial learning of rats by using T-maze task have been investigated.
MATERIALS AND METHODS
Subjects: This study was done in lob of learning and memory, Shahid Chamran university for 4 months in 2006.
So, 50 adult male Wistar rats weighing 275+25 g were used. The rats were housed in individual cages with free access to water and Pellets food. Their ages were 1.5-2 months at the training test.
Group treatment and diets: Palmitic acid (10 g) and standard food (90 g) were mixed then rats were fed freely all the time. Briefly, the diets contained the following: 5% soyabean protein isolate, 0.3% DL-methioninm, 32.7% corn starch, 25% sucrose, 2% cellulose powder, 5% mineral mixture, 1% vitamin mixture and 10% palmitic acid. Rats were categorized as 5 groups (N = 10) as follow:
| Group1 | : | Fed palmitic acid 10% for 1 week |
| Group 2 | : | Fed palmitic acid 10% for 2 weeks |
| Group3 | : | Fed palmitic acid 10% for 3 weeks |
| Group 4 | : | Fed palmitic acid 10% for 4 weeks |
| Group5 | : | As control group was fed ordinary food (Hoseinzadeh et al., 2006) |
Behavioral testing: Palmitic acid (C16H32O2) were obtained from Merck chemical company. All rats were trained in T-maze during 9 days based on standard method (Annet et al., 1989).
Three days before the behavioral test, rats were fasted for 23 h day-1 to maintain 80% of normal body weight. Water was available ad libitum throughout all experiments (Cocco et al., 2002).
Apparatus: A wooden T-maze with 15 cm high walls and start and goal boxes 15Ã-20 cm was used. The 48 cm long stem led to two L-shaped arms. The first part of each arm was 39 cm long and the second part, which led to the goal box, 28 cm long. Guillotine doors separated the start box from the stem and the stem from the L-shaped arms.
Procedure: Preliminary training took place on days 1 to 3. On day 1 food pellets (4 mg) were left in the stem, arms and goal boxes with all the guillotine doors open. The rats were placed individually in the start box and left for 5 min to eat the pellets and explore the maze.
On days 2 and 3 each rat was confined to the start box for 10 sec and then allowed to find 4 pellets in either of the goal boxes, where it was confined for a further 40 sec.
On days 4 and 5, two pellets were available in both of the goal boxes on every trial. Each rat was confined to the start box for 10 sec before being allowed to choose one of the goal boxes, where it was confined for a further 20 s. Eleven consecutive trials were given per day and the choice of right or left goal box was recorded, as was the latency between leaving the start box and entering the chosen arm.
On days 6, 7 and 8 were considered for spatial learning and reversal. Only one of the goal boxes was now rewarded and the rats had to learn which was correct. For half the rats the reward was to be found in the right goal box and for the other half the left goal box was rewarded. Trials continued until the criterion of 5 consecutive correct responses had been achieved. The choice of arm and latency between leaving the start box and entering the chosen arm were recorded on every trial. Immediately after reaching criterion the contingencies were reversed so that the previously unreinforced goal box was now correct. Training continued until the new response had been learnt, again to a criterion of 5 consecutive correct responses. This procedure commenced on day 6 and was repeated on days 7 and 8. Each day the rat was always required to start by re-learning the spatial discrimination which had been correct last during the previous day's training.
On day 9 as a extinction, after a spatial learning had been completed to 5 consecutive Correct responses, an extinction stage were introduced. The food pellets were removed from the T-maze and goal box choices and start box to arm latencies were recorded over a further 10 trials.
Statistical analysis: For comparison between each ditary group and control were made using student t-test and for compare all groups, then data elevated by two way ANOVA. For comparison two by two ditary groups were assessed by post hot the tukey test.
RESULTS
The result showed that a decrease in trail to criterion simple learning between palmitic acid (10%) fed group for one week were significant (p<0.05) but not another groups. The number of trail to criterion on revers learning, in palmitic acid(10%) fed group (1, 2 weeks) decreased (p<0.01) (Fig. 1). But did not differ in number of error on simple learning compare with control group. A decrease in number of error on revers learning in palmitic acid(10%) fed group for 1 week were significant (p<0.001) but not about the other groups (2, 3 and 4 weeks) (Fig. 2). Mean latencies on simple learning in all of groups were not significant but mean latencies on revers learning and extinction trails in palmitic acid (10%) fed groups for 1, 2, 3 weeks were decreased (p<0.05, p<0.001) (Fig. 3 and 4). Generally, learning parameters in groups fed palmitic acid for 1, 2 and 3 weeks were significantly increased.
According these result and in comparison palmitic acid (10%) fed group for 1, 2, 3 and 4 weeks were assessed by tukey test, in conclusion that if usage of palmitic acid increases, spatial learning will decrease, so the low dosage of palmitic acid has positive effect on spatial learning and high dosage of that probably has negative effect.
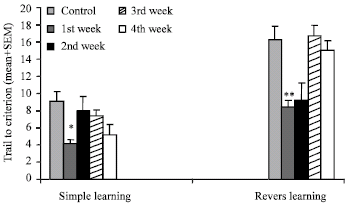 | |
| Fig. 1: | Mean of trail to criterion on simple and revers learning in rat which were fed with a diet containing palmitic acid (10%) 1, 2, 3 and 4 week groups and control, p<0.05*, p<0.01** |
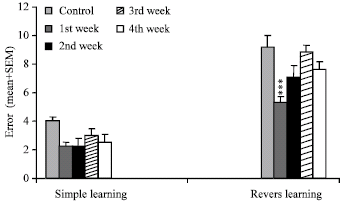 | |
| Fig. 2: | Number of error on simple and revers learning in rat which were fed with a diet containing palmitic acid (10%) 1, 2, 3 and 4 weeks groups and control, p<0.001*** |
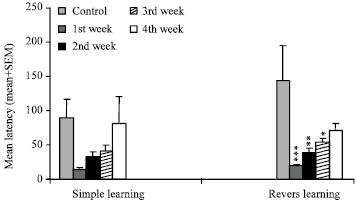 | |
| Fig. 3: | Mean latencies on simple and revers learning in rat which were fed with a diet containing palmitic acid (10%) 1, 2, 3 and 4 weeks groups and control, p<0.05*, p<0.01**, p<0.001*** |
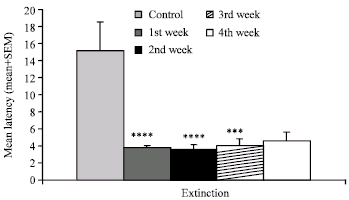 | |
| Fig. 4: | Mean latencies on extinction in rat which were fed with a diet containing palmitic acid (10%)1, 2, 3 and 4 weeks groups and control, p<0.001***, p<0.000**** |
DISCUSSION
The present results point to dietary of palmitic acid (10%) in 1, 2, 3 weeks were significantly increased in spatial learning.
In past studies results showed that the group were dosed lovstatin for 9 days after 2 weeks administration of butter (10%) enhanced learning performance compared to rats that dosed only with lovstatin and historical results studies exhibited that have not had obvious signs of cell injury in rats hippocampus. However increases learning performance that may be due to presence of saturated fatty acids in butter and also the most amount of saturated fatty acids in the butter is palmitic acid. Indeed the developing brain produces all required palmitic acid (Marbios et al., 1992).
Recently investigation show that palmitic modification of neuronal proteins is critical. Protein palmitoylation represents a common lipid modification of neuronal proteins (El-Husseini Ael et al., 2002a; Greaves and Chamberlain, 2007). This posttranslational change involves addition of the saturated 16 carbon palmitic lipid in a thioester linkage to specific cyctein residues. Palmitoylation modifies nurmous important neuronal protein GAP-43, G protein and etc. Importantly, palmitoylation occurs in reversible fashion which allows palmitoylation to dynamically regulation protein function and to participate in diverse aspects of neuronal signaling (Fukata et al., 2004). Moreover, Palmitate modifies both peripheral and integral membrane proteins and its addition can be permanent or transient, which makes it unique among the lipid modifications of proteins. The presence of palmitate on a protein affects how the protein interacts with lipids and proteins in a membrane compartment and the reversibility of palmitoylation allows different modes of trafficking between membrane compartments (Linder and Descheres, 2007). It seems palmitic acid via some mechanisms effects on nerve system and cognition function which here indicated them.
At first, Glutamate receptor channels mediate most of the fast excitory synaptic transmission in the central nervous system. At excitory synapses, ionotropic glutamate receptors such as α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA)/kinate type and N-methyl-D-aspartate (NMDA) type are highly concentrated in the postsynaptic density (PSD) and these receptors are important in learning mechanism. It has recently been proposed that protein-protein interaction of some cytosolic proteins with the C-termini of glutamate receptor subunits are involved in anchoring/clustering receptors at the PSD and coupling the receptors to cytoplasmic signaling molecules (Yamazaki et al., 2001). The postsynaptic density protein, PSD, represents a major palmitoylated protein in brain (Topinka and Bredt, 1998). This lipid modification is critical for PSD-95 clustering of AMPA receptors at excitory synapses. Furthermore, the palmitoylation of PSD-95 is dynamically regulated by synaptic, such that cycling of palmitate on PSD-95 can contribute to aspects of synaptic plasticity and palmitate cyciling on PSD-95 is augmented by calcium influx through NMDA receptors (El-Husseini Ael et al., 2002b). According these study and present results dietary of palmitic acid may be by effects on NMDA receptor induced learning.
On the other hand, Changes in glutamate receptor number at the PSD provide a critical mechanism for rapidly altering synaptic strength (Malawian et al., 2000). Interestingly, the two principle classes of ionotropic glutamate receptors are differentially anchored at the PSD. NMDA receptors are firmly tethered to the PSD, whereas the AMPA receptors are more loosely attached (Malawian et al., 2000; Malenka and Nicoll, 1999). Dynamic cycling of AMPA receptors on and off the synaptic membrane occurs in an activity-dependent fashion and appears to synaptic plasticity (Beattie et al., 2000) furthermore, the PDZ domains from the PSD-95 were first shown to bind the c-terminal tails of NMDA receptor typ-2 subunit and shaker-type k+channels (Kim et al., 1995). However, the PSD-95 family of membrane-associated guanylate kinase (MAGUK) can also interact with AMPA receptors. Furthermore, the AMPA receptor trafficking protein, stargazing, associates with PDZ domains from several neuronal MAGUKS (Chen et al., 2000) and MAGUKS proteins regulate synapse development and plasticity. In cultured hippocampal neurons, overexpression of PSD-95 accelerates development of excitory synapses and selectively enhances clustering of AMPA receptors (El-Husseini Ale et al., 2000). On the other hand, targeted disruption of PSD-95 causes sever abnormalities of synaptic plasticity, such as long-term potentiation (LTP) enhanced and long-term depression(LTD) is eliminated. These abnormalities in synaptic plasticity presumably explain why PSD-95 mutant mice are impaired in spatial learning (Magaud et al., 1998). It verifies these results about positive effect of palmitic acid dietary in 1, 2 and 3 weeks.
Also, Nerve Growth Factor (NGF) is critical for neuronal survival and for the establishment and maintenance of synaptic connection (Chao, 2003). NGF binds the tropomyosin-related kinase receptor A (TrkA) and P75 receptor with comparable affinities. Indeed Ncam1 is the best-characterized member of the calcium independent adhesion molecules (Bartsch, 2003).
It is present throughout the developing nervous system and its expression persist in the adult (Montag-Sallaz et al., 2003). During brain development, Ncam1 is heavily polysialylated, while the developing brain has a high level of Ncam1, the adult brain has appreciable levels only in the areas that continually undergo structural reorganization and demonstrate synaptic plasticity, such as the olfactory bulb, the hypothalamus and hippocampus (Bukalo et al., 2004). Ncam1 is transiently upregulated at the synapse during learning and memory (Sytnyk et al., 2004). So, it may be dietary of palmitic acid via regulation this molecule effect on spatial learning because palmitic acid can cross from blood barrier brian.
Moreover, P75 receptor is required for both resting and NGF-induced Ncam1 expression. NGF-induced Ncam1 expression is dependent on the presence of an intact palmitoylation site within P75 receptor (Mirnics et al., 2005). It may be palmitic acid by regulate this factors effects on learning function. The gamma 2 subunit of neuronal and recombinant GABA receptors is palmitoylation. In cultured hippocampal neurons, inhibitors of protein palmitoylation reduced the synaptic clusturing of GABA receptors and steady-state cell surface receptor number (Ruthenberg et al., 2004), so palmitoylation is critical for regulation GABA receptor. Interestingly an increased impulse flow in the septohippocampal GABAergic pathway facilitated enhanced cognition initiated by a muscarinic agonist (Li et al., 2002).
GABAergic neurons sense the excitory transmission and regulate synaptic strength by sending feed forward and/or feedback inhibitory inputs to the principal neurons. Regulation of synaptic efficacy by integrated excitory and inhibitory transmission within specific neuronal network is thought to underlie memory encoding and retrieval in hippocampus (Zhao et al., 2004). Present of palmitic acid is necessary for regulation molecular mechanism that influence in learning and memory which prove these result about positive effect of dietary palmitic acid at 1, 2 and 3 weeks on spatial learning (Fig. 1-4).
On the other hands, Epidemiological studies suggest that high fat diets significantly increase the risk of Alzheimer's Disease (AD). In addition, the AD brain is characterized by high fatty acid content compared to that of healthy subjects. Nevertheless, the basic mechanism relating elevated fatty acids and the pathogenesis of AD remains unclear. The present study examines the role of fatty acids in causing hyperphosphorylation of the tau protein, one of the characteristic signatures of AD pathology. Hyperphosphorylation of tau disrupts the cell cytoskeleton and leads to neuronal degeneration (Patil et al., 2005). Present result indicted spatial learning in a group fed palmitic acid for 4 week was not only increased but also it is decreased.
CONCLUSION
According these studies regulate many protein in nerve system by palmitoylation is very important in cognition process such as, learning and memory.
ACKNOWLEDGMENT
The author express their gratitude to the research council of Shahid Chamran University for their financial supports.
REFERENCES
- Bartsch, U., 2003. Neural CAMS and their role in the development and organization of myelin sheaths. Front. Biosci., 8: d477-d490.
PubMedDirect Link - Bukalo, O., N. Fentrop and A.Y. Lee, 2004. Conditional ablation of the neural cell adhesion molecule reduces precision of spatial learning long-term potentiation and depression in CA1 subfield of mouse hippocampus. J. Neurosci., 24: 1565-1577.
CrossRefDirect Link - Chao, M.V., 2003. Neurotorophins and their receptors: A convergence point for many signaling pathways. Neurosci., 4: 299-309.
CrossRefDirect Link - Chen, L., D.M. Chetkovich and R. Petraillia, 2000. Stargazin mediates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature, 408: 936-943.
CrossRefDirect Link - Cocco, S., G. Diaz, R. Stancampiano, A. Diana and M. Carta et al., 2002. Vitamin A deficiency produces a spatial learning and memory impairment in rats. Neuroscience, 115: 475-482.
Direct Link - Edmond, J., T.A. Higa and R.A. Korsak, 1998. Fatty acid transport and utilization for the developing brain. J. Neurochem., 70: 1227-1234.
PubMedDirect Link - El-Husseini Ael, D. and D.S. Bredt, 2002. Protein palmitoylation:A regulator of neuronal development and function. Nat. Neurosci., 3: 791-802.
PubMedDirect Link - El-Husseini, A.D., E. Schnell, S. Dakoji, N. Sweeney and Q. Zhou et al., 2002. Synaptic strength regulated by palmitate cycling on PSD-95. Cell, 108: 849-863.
CrossRefDirect Link - Fukata, M., Y. Fukata, H. Adesnik, R.A. Nicoll and D.S. Bredt, 2004. Identification of PSD-95 palmitoylating enzymes. Neuron, 44: 987-996.
CrossRefDirect Link - Greaves, J. and L.H. Chamberlain, 2007. Palmitoylation-dependent protein sorting. J. Cell Biol., 176: 249-254.
CrossRefPubMedDirect Link - Kodas, E., L. Galineau and S. Bodard, 2004. Serotoninergic neurotransmission is affected by n-3 polyunsaturated fatty acids in the rat. J. Neurochem., 89: 695-702.
PubMedDirect Link - Laasonen, M., A.T. Erkkila, E. Isotalo and P.K. Maenpa, 2006. Serum lipid fatty acids and temporal processing acuity in children with oral clefts. Prostaglandins, Leukotrienes Essential Fatty Acids, 74: 263-270.
PubMedDirect Link - Li, S., M. Park and J. Bahk, 2002. Chronic nicotine and smoking exposure decreases GABAB1 receptor expression in the rat hippocampus. Neuroscience, 334: 135-139.
CrossRefDirect Link - Linder, M.E., R.J. Deschenes, 2007. Palmitoylation: Policing protein stability and traffic. Nat. Rev. Mol. Cell Biol., 8: 74-84.
PubMedDirect Link - Magaud, M., P. Charlesworth and M. Dempster, 1998. Enhanced long-term potentiation and impaired learning in mice with mutant postsynaptic density-95 protein. Nature, 396: 433-439.
CrossRefDirect Link - Malenka, R.C., R.A. Nicoll, 1999. Role of AMPA receptor cycling in synaptic transmission and plasticity. Neuron, 24: 649-658.
PubMedDirect Link - Malawian, R., Z.F. Mainen and Y. Hayashi, 2000. LTP mechanisms: From silence to four-lane traffic. Curr. Opin. Neurobiol., 10: 352-357.
CrossRefDirect Link - Marbios, B.N., H.O. Ajie, R.A. Korsak, D.K. Sensharma and J. Edmond, 1992. The effect of palmitic acid in brain of the developing rat. Lipid., 27: 587-592.
Direct Link - McCann, J.C. and B.N. Ames, 2007. Is docosahexaenoic acid, an n_3 long-chain polyunsaturated fattyacid, required for development of normal brain function? An overview of evidence from cognitive and behavioral tests in humans and animals. Am. J. Clin. Nutr., 82: 281-295.
Direct Link - Mirnics, Z.K., C. Yan and C. Portugal, 2005. P75 neurotrophin receptor regulation expression of neural cell adhesion molecule 1. Neurobiol. Dis., 20: 969-985.
PubMedDirect Link - Montag-Sallaz, M., D. Montag and M. Schachner, 2003. Altered processing of novel information in N-CAM-deficient mice. Neuro Rep., 14: 1343-1346.
PubMedDirect Link - Ruthenberg, J., J.T. Kittler and S.J. Moss, 2004. Palmitoylation regulates the clustering and cell surface stability of GABAA receptors. Mol. Cell Neurosci., 26: 251-257.
PubMedDirect Link - Ruthrich, H., G. Grecksch and M. Krug, 1999. Effects of piracetam on pentyleneterazol-kindling development, hippocapal potentiation phenomena and kindling-induced learning deficit. Naunyn Schmiedebergs Arch. Pharmacol., 360: 413-420.
Direct Link - Sytnyk, V., I. Leshchynska and A. Dityatev, 2004. Transgolgi network delivery of synaptic proteins in synaptogenesis. J. Cell Sci., 117: 381-388.
CrossRefDirect Link - Yehuda, S., 2003. Omega-6/omega-3 ratio and brain-related function. Sci. Evidence, 92: 37-56.
Direct Link - Yehuda, S., S. Rabinovitz and D.I. Mostofsky, 2006. Nutritional deficiencies in learning and cognition. J. Pediatr. Gastroenterol. Nutr. Des., 43: S22-S25.
CrossRefDirect Link - Yamazaki, M., M. Fukaya and M. Abe, 2001. Differential palmitoylation of two mouse glutamate receptor interacting protein 1 forms with different N-terminal sequences. Neurosci. Lett., 304: 81-84.
CrossRefDirect Link - Zhao, W., H. Chen and M.J. Quon, 2004. Insulin and the insulin receptor in experimental models of learning and memory. Eur. J. Pharmacol., 490: 71-81.
PubMedDirect Link