
Nattapong Srisamoot
Department of Biochemistry
Arunrat Chaveerach
Department of Biology, Faculty of Science, Khan Kaen University, Khan Kaen, 40002, Thailand
Suporn Nuchadomrong
Department of Biochemistry
Nison Sattayasai
Department of Biochemistry
Prapansak Chaveerach
Department of Veterinary Public Health, Faculty of Veterinary Medicine,
Khan Kaen University, Khan Kaen, 40002, Thailand
Alongkoad Tanomtong
Department of Biology, Faculty of Science, Khan Kaen University, Khan Kaen, 40002, Thailand
Krit Pinthong
Biology Program, Faculty of Science and Technology, Surin Rajabhat University, Surin, 32000, Thailand
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 16 | Page No.: 2639-2645







ABSTRACT
The cytogenetics of eight Felidae species in Thailand were investigated by the colchicines-hypotonic fixation-air drying technique followed by a conventional technique. All species studied have an identical number of 38 diploid chromosomes, indicating a close genetic relationship among species. At a deep study level, the genetic relationships of eight Felidae species were accessed by the AFLP method. Blood samples were collected from sources locating in their original regions for DNA extraction. With ten successful primer combinations, a total of 4208 scorable bands were generated. Of these bands, 18.91% are polymorphic. Percentages of Polymorphic Bands (PPB) for each primer combination range from 15.00 to 23.59%. The generating bands were used for dendrogram construction. The average genetic similarity values among all Felidae species are 68.20% (between Panthera tigris and Neofelis nebulosa) to 85.53% (between Prionailurus bengalensis and Prionailurus viverrinus). The dendrogram shows that the eight Felidae species were clustered together and the subfamily Pantherinae and Felinae with Neofelis nebulosa are distinguished. The Felinae, Prionailurus bengalensis, Prionailurus viverrinus, Catopuma temminckii, Felis chaus, Pardofelis marmorata and Neofelis nebulosa were clustered together with 91% bootstrap support and the Pantherinae, Panthera pardus is clustered with Panthera tigris with 92% bootstrap support. In summary, the ten successful primer combinations can be used to determine genetic differences among eight Thailand Felidae species.
PDF Abstract XML References Citation
How to cite this article
Nattapong Srisamoot, Arunrat Chaveerach, Suporn Nuchadomrong, Nison Sattayasai, Prapansak Chaveerach, Alongkoad Tanomtong and Krit Pinthong, 2007. Genetic Relationships among Wild Felidae in Thailand Using AFLP Markers. Pakistan Journal of Biological Sciences, 10: 2639-2645.
DOI: 10.3923/pjbs.2007.2639.2645
URL: https://scialert.net/abstract/?doi=pjbs.2007.2639.2645
DOI: 10.3923/pjbs.2007.2639.2645
URL: https://scialert.net/abstract/?doi=pjbs.2007.2639.2645
INTRODUCTION
All members of Felidae were considered from major population in Appendix I and II of CITES and on the IUCN Red list of threatened, endangered and extinct species (Baillie et al., 2004). Thailand has many charismatic felid species in the world, including tiger (Panthera tigris), leopard (Panthera pardus), clouded leopard (Neofelis nebulosa), asian leopard cat (Prionailurus bengalensis), fishing cat (Prionailurus viverrinus), flat-headed cat (Prionailurus planiceps), asian golden cat (Catopuma temminckii), jungle cat (Felis chaus) and marbled cat (Pardofelis marmorata). Nine species of Felidae in Thailand can be divided into two subfamilies, Felinae and Pantherinae, based on morphological classification (Wilson and Reeder, 2006). Habitat loss has occurred violently throughout Southeast Asia over the past 20 years and these felid species are vulnerable to population pressures and habitat fragmentation. Forest destruction has negatively affected on wild animals such as Felidae. It reduces habitat for wild animals and causes population fragmentation due to the loss of genetic heterogeneity and thus they become vulnerable to environmental change and risk extinction.
Molecular genetic data for Felidae in Thailand, such as genetic relationships, is directly needed for conservation. The phylogenetic relationships among the Felidae have been addressed by several studies that employed both morphological and molecular techniques. Early efforts included comparative karyology (Modi and O’Brien, 1988; Wurster-Hill and Centerwall, 1982), cross-species chromosome painting (Tian et al., 2004), the genomic occurrence of two felid endogenous retroviruses (Benveniste and Todaro, 1974; Benveniste et al., 1975; Reeves and O’Brien, 1984), albumin immunological distance (Collier and O’Brien, 1985), comparative morphology (Salles, 1992), allozyme electrophoresis (O’Brien et al., 1987; Pecon-Slattery et al., 1994) and two-dimensional protein electrophoresis (Pecon-Slattery et al., 1994). More recently, efforts to resolve phylogenetic relationships have focused on the nuclear DNA sequence (Flynn et al., 2005; Sato et al., 2006; Zhang et al., 2006) and mitochondrial genome (Janczewski et al., 1995; Johnson et al., 1996; Lopez et al., 1994; Uphyrkina et al., 2002; Johnson et al., 2004; Flynn et al., 2005; Koepfli et al., 2006). However, few analyses have included information from Amplified Fragment-Length Polymorphism (AFLP).
AFLP has become a popular marker technique in genetic relationships because it combines restriction digestion and amplification of DNA fragments, with no prior knowledge about the target genome required (Vos et al., 1995). Abundant polymorphism and reproducibility have been proved to be advantages of the AFLP technique (Pejic et al., 1998; Powell et al., 1996; Russell et al., 1997). The high frequency of identifiable polymorphic AFLP markers, coupled with their reproducibility, make this technique an attractive tool for detecting polymorphism and determining genetic relationships (Gupta et al., 1999). Moreover, the genetic relationships within Felidae in Thailand have never been addressed using AFLP analysis.
In this study we investigated the cytogenetics data and AFLP was applied to estimate genetic relationships of eight Felidae species in Thailand, without Prionailurus planiceps because of lack of sample.
MATERIALS AND METHODS
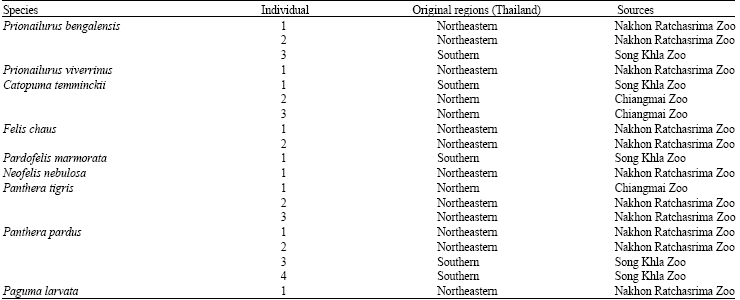
Sample collection: Blood samples of Prionailurus bengalensis, Prionailurus viverrinus, Catopuma temminckii, Felis chaus, Pardofelis marmorata, Neofelis nebulosa, Panthera tigris and Panthera pardus were taken from sources locating in their original regions for DNA extraction as shown in Table 1 in 2003-2004. AFLP fingerprints were done at Department of Biochemistry, Faculty of Science, Khon Kaen University, Khon Kaen, Thailand in 2004-2006. As both morphological and molecular evidence place the family Viverridae as a successive sister-group to the Felidae (Mattern and McLennan, 2000), so the Viverridae, Paguma larvata (Masked-palm civet) was chosen as an outgroup in our analysis.
Cytogenetic analysis: The cultured cells were examined by the colchicines-hypotonic fixation-air drying technique followed by a conventional technique (Tanomtong et al., 2005). Chromosomal checks were performed with 20 cells of each individual by light microscopy.
DNA extraction: Genomic DNA was isolated from blood samples using protinase K digestion and treatment with phenol/chloroform (Sambrook et al., 1989). The quality and quantity of extracted DNA was checked by 0.8% agarose gel electrophoresis and spectrophotometry.
AFLP procedure: The procedure of the AFLP method (Vos et al., 1995) was performed according to the protocol of the kit (AFLPR Analysis System I, Invitrogen, USA).
| Table 1: | Lists of individual number of each species collected from different original regions and sources |
 | |
After adaptor ligation and preselective amplification, selective amplification was conducted with 24 primer combinations and 10 primer combinations: EcoRI-AAC/MseI-CAA, EcoRI-AAC/MseI-CTG, EcoRI-AAC/MseI-CTT, EcoRI-AAG/MseI-CAA, EcoRI-ACA/ MseI-CAG, EcoRI-ACA/ MseI-CAT, EcoRI-ACA/ MseI-CTA, EcoRI-ACA/ MseI-CTC, EcoRI-ACA/ MseI-CTG and EcoRI-ACA/ MseI-CTT were successful. The PCR products amplified with different primer combinations were loaded onto 6.0% denaturing polyacrylamide gels and electrophoresed for 3 h and detected by Silver QuestTM Silver Staining Kit (Invitrogen, USA).
Data analysis: The AFLP bands were visually scored as either present (1) or absent (0) for each accession and each primer combination without band intensity consideration. With the 0/1 data, a pair-wise genetic distance value was generated using Pearson correlation coefficients and these values were then converted to a genetic distance matrix. Based on the genetic distance matrix, cluster analyses were performed and the corresponding dendrogram was constructed for Felidae using arithmetic means analysis (UPGMA). Unbiased measures of genetic similarity index (S) were also calculated between each pair of the species. All these analyses were done by the FingerprintingTM II Software (BIO-RAD, USA).
RESULTS
Results from lymphocyte culture of whole blood and conventional staining of the eight Felidae species, namely Prionailurus bengalensis, Prionailurus viverrinus, Catopuma temminckii, Felis chaus, Pardofelis marmorata, Neofelis nebulosa, Panthera tigris and Panthera pardus indicate that all species have an identical number of 38 diploid chromosomes (2n) consisting of 36 autosomes and 2 sex chromosomes. A representative chromosome of the eight Felidae species from a male of Prionailurus bengalensis is shown in Fig. 1.
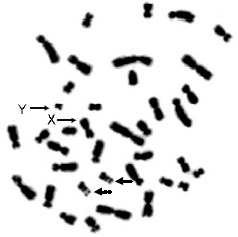 | |
| Fig. 1: | Diploid chromosome number of male Prinailurus bengalensis, 2n = 38 consisting of 36 autosomes and 2 sex chromosomes. The satellite chromosomes are indicated bv dash arrows |
A total of 4208 scorable bands, with sizes ranging from approximately 100-1500 base pairs (bp), were generated using ten EcoRI/MseI AFLP primer combinations with the entire collection of 18 Felidae individuals (Table 2). The number of scorable bands for each primer combination ranged from 320 to 518 (mean 420.8). Of these bands, 18.91% are polymorphic. Percentages of Polymorphic bands for each primer combination range from 15.00 to 23.59%. The most informative primer combination is the EcoRI-ACA/MseI-CTC pair, which produced the highest number of bands (518), while primer EcoRI-AAC/MseI-CTG combinations produced a minimum number of bands (320). Representative AFLP profiles of 18 Felidae individuals generated by the EcoRI-ACA/MseI-CTG primer combination are shown in Fig. 2.
The dendrogram constructed by the UPGMA method demonstrate that the 8 Felidae species are clustered together and the subfamilies Pantherinae and Felinae can be separated into two groups, whereas the outgroup Paguma larvata is segregated from the Felidae with a high bootstrap value (Fig. 3). In subfamily Felinae, Prionailurus bengalensis, Prionailurus viverrinus, Catopuma temminckii, Felis chaus and Pardofelis marmorata are clustered together with 91% bootstrap support attaching with Neofelis nebulosa of Pantherinae.
| Table 2: | A summary of AFLP primers, number of bands scored, number of polymorphic bands and percentages of polymorphisms for amplification profiles of eight felidae species in Thailand |
 | |
 | |
| Fig. 2: | Banding patterns of AFLP analysis among 18 Felidae accessions generated by the EcoRI-ACA/MseI-CTG primer combination |
 | |
| Fig. 3: | Dendrogram of eight Felidae species constructed based on UPGMA clustering |
In subfamily Pantherinae, Panthera pardus is clustered with Panthera tigris without Neofelis nebulosa with 92% bootstrap support. Moreover, Neofelis nebulosa produces a sister species to the Felinae group with high bootstrap value (90%).
| Table 3: | The average similarity index values based on the AFLP analysis of the 8 felidae species and Masked-palm civet |
 | |
The average genetic similarity index (S) of 8 species ranged from 68.20% between Panthera tigris and Neofelis nebulosa to 85.53% between Prionailurus bengalensis and Prionailurus viverrinus (Table 3).
DISCUSSION
In the present study, the cytogenetic data demonstrate that eight Felidae species have an identical diploid chromosome number (2n = 38), agreeing with Hsu and Rearden (1965), Wurster-Hill and Benirschke (1968), Wurster-Hill (1969), Wurster-Hill and Gray (1973) and Pathak and Wurster-Hill (1977) and reflecting the close genetic relationships among species of Felidae. However, the chromosome number does not provide deep information of determination genetic relationships among Felidae, so the AFLP molecular marker was used to elucidate. AFLP markers were successfully used to survey genetic variation and relationships among eight species of Felidae in Thailand. The ability to determine genetic variation among species at molecular level is directly related to the number of polymorphisms detected and their reproducibility. From resulting bands from ten primer combinations, a low level of polymorphism was found among Felidae species agreeing with the identical chromosome number. Prionailurus viverrinus is very closely related to Prionailurus bengalensis based on the highest S value, 85.53%, while the lowest S value is between Panthera tigris and Neofelis nebulosa. The S values between individuals in a species were generally high (>80%) (data not shown).
The dendrogram shows that 8 species of Felidae are separated into two groups, subfamilies Pantherinae and Felinae which includes Neofelis nebulosa possessing a relatively high S value to Felinae species than to other Pantherinae, which is contrary to morphological classification, but agree with the shape and size according to Felinae. The separation of Felidae into two groups of big cats (Pantherinae) and small cats (Felinae) is generally supported by most studies. A clear size dichotomy does seem to exist in Felidae (Bininda-Emonds et al., 2001). Neofelis nebulosa which is the smallest of the big cats with a body size of 16-23 kg (Mattern and McLennan, 2000) and the shape of a small cat has teeth and skull structure which is clearly that of a Pantherinae (Postanowicz, 2006). Support for monophyletic groups composed of Pantherinae species is abundant from morphological characters (Neff, 1982; Peters and Hast, 1994; Salles, 1992).
Based on present study, we found a close relationship between Pardofelis marmorata and Neofelis nebulosa with high S value according to the phylogenetic tree recently constructed by Bininda-Emonds et al. (1999) and Ortolani (1999). The dendrogram shows that Catopuma temminckii has a stronger genetic relationship to Prionailurus bengalensis and Prionailurus viverrinus than Felis chaus, while Bininda-Emonds et al. (1999), Mattern and McLennan (2000) and Pecon-Slattery et al. (2004) reported that Felis chaus is more closely related to Prionailurus bengalensis and Prionailurus viverrinus than Catopuma temminckii. Present results agreeing with the phylogenetic relationships of Felidae based on analyses of combination 16S rRNA with NADH-5 mtDNA gene segments by Johnson and O’Brien (1997) and the phylogeny of body and tail color pattern, tail tip markings, eye contour and eye patches constructed by Ortolani (1999) shows that Catopuma temminckii comprises a sister species to Prionailurus bengalensis and Prionailurus viverrinus, while Felis chaus forms a sister species to these groups. However, Prionailurus bengalensis and Prionailurus viverrinus are classified to the Asian leopard cat group whereas Catopuma temminckii, only one species in Thailand, is classified to the Bay cat group (Johnson and O’Brien, 1997). In order to resolve the genetic relationships between the Asian leopard cat group and Catopuma temminckii properly, it is necessary to study a larger sample. However, large sample studies of wildlife are most difficult, causing the small sample number studied in our research, so that the method for producing reproducible bands and abundant polymorphisms like AFLP has been used. Thus, these results suggest that genetic analysis based on the AFLP technique has a good capacity for the study of genetic relationships, especially in Felidae. As AFLP analysis does not require prior genetic information for the taxa, this technique should be of value in genetic analysis of wild Felidae. Moreover, the ten successful primer combinations can be used to determine genetic differences among the eight Felidae species in Thailand.
ACKNOWLEDGMENTS
This study was supported by research fund from Thailand Zoological Park Organization under the Royal Patronage of His Majesty the King.
REFERENCES
- Bininda-Emonds, O.R.P., J.L. Gittleman and A. Purvis, 1999. Building large trees by combining phylogenetic information: a complete phylogeny of the extant Carnivora (Mammalia). Biol. Rev., 74: 143-175.
Direct Link - Bininda-Emonds, O.R.P., D.M. Decker-Flum and J.L. Gittleman, 2001. The utility of chemical signals as phylogenetic characters: An example from the felidae. Biol. J. Linn. Soc., 72: 1-15.
Direct Link - Collier, G.E. and S.J. O`Brien, 1985. A molecular phylogeny of the felidae: Immunological distance. Evology, 39: 473-487.
Direct Link - Flynn, J.J., J.A. Finarelli, S. Zehr, J. Hsu and M.A. Nedbal, 2005. Molecular phylogeny of the Carnivora (Mammalia): Assessing the impact of increased sampling on resolving enigmatic relationships. Syst. Biol., 54: 317-337.
CrossRef - Gupta, P.K., R.K. Varshney, P.C. Sharma and B. Ramesh, 1999. Molecular markers and their applications in wheat breeding. Plant Breed., 118: 369-390.
CrossRefDirect Link - Janczewski, D.N., W.S. Modi, J.C. Stephens and S.J. O`Brien, 1995. Molecular evolution of mitochondrial 12S RNA and cytochrome b sequences in the pantherine lineage of felidae. Mol. Biol. Evol., 12: 690-707.
Direct Link - Johnson, W.E., P.A. Dratch, J.S. Martenson and S.J. O`Brien, 1996. Resolution of recent radiations within three evolutionary lineages of felidae using mitochondrial restriction fragment length polymorphism variation. J. Mammal. Evol., 3: 97-120.
CrossRef - Johnson, W.E. and S.J. O’Brien, 1997. Phylogenetic reconstruction of the Felidae using 16SrRNA and NADH-5 mitochondrial genes. J. Mol. Evol., 44: S98-S106.
CrossRefDirect Link - Johnson, W.E., J.A. Godoy, F. Palomares, M. Delibes, M. Fernandes, E. Revilla and S.J. O’Brien, 2004. Phylogenetic and phylogeographic analysis of iberian lynx populations. J. Hered., 95: 19-28.
Direct Link - Koepfli, K., S.M. Jenks, E. Eizirik, T. Zahirpour, B.V. Valkenburgh and R.K. Wayne, 2006. Molecular systematics of the hyaenidae: Relationships of a relictual lineage resolved by a molecular supermatrix. Mol. Phylogenet. Evol., 38: 603-620.
Direct Link - Lopez, J.V., N. Yuhki, R. Masuda, W. Modi and S.J. O'Brien, 1994. Numt, A recent transfer and tandem amplification of mitochondrial DNA to the nuclear genome of the domestic cat. J. Mol. Evol., 39: 174-190.
CrossRefDirect Link - Mattern, Y.M. and A.D. McLennan, 2000. Phylogeny and speciation of felids. Cladistics, 16: 232-253.
CrossRef - Ortolani, A., 1999. Spots, stripes, tail tips and dark eyes: Predicting the function of carnivore colour patterns using the comparative method. Biol. J. Linn. Soc., 67: 433-476.
CrossRef - Pecon-Slattery, J., W.E. Johnson, D. Goldman and S.J. O’Brien, 1994. Phylogenetic reconstruction of South American felids defined by protein electrophoresis. J. Mol. Evol., 39: 296-305.
CrossRefDirect Link - Pecon-Slattery, J., A.J. Pearks Wilkerson, W.J. Murphy and S.J. O’Brien, 2004. Phylogenetic assessment of introns and SINEs within the Y chromosome using the cat family felidae as a species tree. Mol. Biol. Evol., 21: 2299-2309.
Direct Link - Pejic, I., P. Ajmone-Marsan, M. Morgante, V. Kozumplick, P. Castiglioni, G. Taramino and M. Motto, 1998. Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs and AFLPs. Theor. Applied Genet., 97: 1248-1255.
CrossRefDirect Link - Peters, G. and M.H. Hast, 1994. Hyoid structure, laryngeal anatomy and vocalization in felids (Mammalia: Carnivora: Felidae). Z. Saugetierkunde, 59: 87-104.
Direct Link - Powell, W., M. Morgante, C. Andre, M. Hanafey, J. Vogel, S. Tingey and A. Rafalski, 1996. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed., 2: 225-238.
CrossRefDirect Link - Russell, J.R., J.D. Fuller, M. Macaulay, B.G. Hatz, A. Jahoor, W. Powell and R. Waugh, 1997. Direct comparison of level of genetic variation among barley accessions detected by RFLPs, AFLPs, SSRs and RAPDs. Theor. Applied Genet., 95: 714-722.
Direct Link - Sambrook, J., E.F. Fritsch and T.A. Maniatis, 1989. Molecular Cloning: A Laboratory Manual. 2nd Edn., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, USA., ISBN-13: 9780879695774, Pages: 397.
Direct Link - Sato, J.J., M. Wolsan, H. Sizuki, T.Hosoda, Y. Yamaguchi, K. Hiyama, M. Kobayashi and S. Minami, 2006. Evidence from nuclear DNA sequences sheds light on the phylogenetic relationships of pinnipedia: Single origin with affinity to musteloidea. Zool. Sci., 23: 125-146.
Direct Link - Tian, Y., W. Nie, J. Wang, M.A. Ferguson-Smith and F. Yang, 2004. Chromosome evolution in bear: Reconstructing phylogenetic relationships by cross-species chromosome painting. Chromosome Res., 12: 55-63.
Direct Link - Uphyrkina, O., D. Miquelle, H. Quigley, C. Driscoll and S.J. O’Brien, 2002. Conservation genetics of far eastern leopard (Panthera pardus orientalis). J. Hered., 93: 303-311.
Direct Link - Vos, P., R. Hogers, M. Bleeker, M. Reijans and T. van de Lee et al., 1995. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res., 23: 4407-4414.
CrossRefPubMedDirect Link - Wurster-Hill, D.H. and W.R. Centerwall, 1982. The interrelationships of chromosome banding patterns in canids, hyena and felids. Cytogenet. Cell. Genet., 34: 178-192.
PubMed - Zhang, W., Z. Zhang, F. Shen, R. Hou, X. Lv and B. Yue, 2006. Highly conserved D-loop-like nuclear mitochondrial sequences (Numts) in tiger (Panthera tigris). J. Genet., 85: 107-116.
CrossRefDirect Link