
Demir KOK
Department of Horticulture, Faculty of Agriculture, Namik Kemal University, Tekirdag, Turkey
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 16 | Page No.: 2631-2638







ABSTRACT
This research was carried out under the laboratory conditions of Department of Horticulture, Agriculture Faculty, Namik Kemal University in Turkey. In this research, salinity tolerance of V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes derived from Marmara Region and Akdeniz Region of Turkey was evaluated for various salinity levels at seed germination and plantlet stages. In addition to V. vinifera subsp. sylvestris (C.C. Gmelin); 5 BB Kober (V. berlandieri Planch. x V. riparia Michx.) and Isabella grape (V. labrusca L.) were also inserted to study to make a comparison. In order to determine responses of V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes, 5 BB Kober and Isabella grape to salinity, NaCl was added at 0 (control), 2.7, 5.4, 8.1 and 10.8 dS m-1 to nutrient solution to achieve five salinity levels. Prior to study, all seeds were extracted from grape berries and stratified to be permeable to water by humidified sand. Afterwards, seeds were germinated under the different salinity stress conditions mentioned above. At the end of germination phases, germination percentages were calculated for all seed types and fresh weight (mg), dry weight (mg), water content (%), tolerance index values, Na+:K+ values were found out for shoots and roots of all plantlet types. No germination was observed during the germinations of all seeds under the stress conditions induced by 10.8 dS m-1 NaCl treatment. On the basis of various salinity tolerance indexes, it was seen that Marmara Region plantlets were more resistant than Akdeniz Region for 8.1 dS m-1 NaCl treatment. In conclusion, since V. vinifera subsp. sylvestris (C.C. Gmelin) ecotype from Marmara Region exhibits higher resistance to salinity, its rooted plant materials as grapevine rootstock can be used for salinity soil conditions in grape growing. Besides, it can be utilized from seeds of Marmara Region to obtain salinity resistant hybridized grapevine rootstocks in breeding programs of viticulture.
PDF Abstract XML References Citation
How to cite this article
Demir KOK, 2007. Responses of V. vinifera subsp. sylvestris (C.C. Gmelin) Ecotypes Originated from Two Different Geographical Regions of Turkey to Salinity Stress at Seed Germination and Plantlet Stages. Pakistan Journal of Biological Sciences, 10: 2631-2638.
DOI: 10.3923/pjbs.2007.2631.2638
URL: https://scialert.net/abstract/?doi=pjbs.2007.2631.2638
DOI: 10.3923/pjbs.2007.2631.2638
URL: https://scialert.net/abstract/?doi=pjbs.2007.2631.2638
INTRODUCTION
Plant productivity is severely affected by abiotic stress factors which include salinity, drought, high and low temperature, and heavy metals. As a sequel to it, physiological and biochemical responses in plants vary and cellular aqueous and ionic equilibriums are disrupted (Sreenivasulu et al., 2006).
Plant development and successful crop production require suitable soil conditions, including adequate water and supply. Unfavorable soil conditions such as environmental stress (Bohnert et al., 1995) salinity and alkalinity (Qadir et al., 1996), inadequate nutrient supply (Qadir et al., 1997), have an adverse effect on the life of the plants, sometimes seriously hindering their effective production. Plant growth in saline soils is affected mainly by the reduced availability of water due to the high osmotic pressure (Orak and Ates, 2005).
Salinity is a major abiotic stress reducing the yield of a wide variety of crops all over the world (Tester and Davenport, 2003; Ashraf and Foolad, 2007).
Saline soils and water shortage severely limit the productivity of crop in semiarid and arid environments. The response of plant to excess sodium chloride (NaCl) is complex and involves change in their morphology, physiology and metabolism (Parida and Das, 2005).
Salinity interacts with certain plant and environmental factors during germination. Among the plant factors, seed coat, dormancy (Khan and Ungar, 1997), seed age (Smith and Dobrenz, 1987), seed polymorphisim (Khan and Ungar, 1984) and seedling vigor (Lin and Kao, 1995) are prominent and temperature (Khan and Ungar, 1997), light (Villiers De et al., 1994), availability of water (Hegarty, 1978) and oxygen (Wijte and Gallagher, 1996) take place in environmental factors.
Improved production efficiency can be achieved by developing stress tolerant crop (Ribaut et al., 1997), which necessitates research on plant adaptation mechanisms against adverse environmental conditions.
In order to develop salt-tolerant crops, it is necessary to identify the degree of salinity tolerance within crops and their wild-type relatives. Investigations of genetically based differences between closely related plants are particularly important because these studies provide necessary information to select traits for breeding salt-tolerant crops (Tester and Davenport, 2003).
Plants differ in their tolerance to salt and fruits, vegetables and ornamentals are generally more salt sensitive than other crops (Maas, 1990). However, own rooted- V. vinifera L. varieties are highly salinity tolerant, usage of grapevine saplings grafted on grapevine rootstocks is preferred to reduce harmful effects of Phylloxera vastatrix Planchon in viticulture.
V. vinifera subsp. sylvestris (C.C. Gmelin) spontaneously grows in not only woodland regions of Turkey but also various woodland regions of the world and exhibits extensively type richness. From this point of view, Turkey has also rich potential and it is possible to come across with different wild grapevine ecotypes depending on geographical regions. After examining other characteristics of V. vinifera subsp. sylvestris (C.C. Gmelin) (e.g., scion and rootstock compatibilities between grape varieties and V. vinifera subsp. sylvestris (C.C. Gmelin) in grafting; qualities and yields of grape varieties grafted on V. vinifera subsp. sylvestris (C.C. Gmelin)), it can be used for grapevine rootstock with salinity resistant characteristic on non-Phylloxera vastatrix Planchon carrying soils.
The main objective of present study was to evaluate whether there was variation between V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes belonging to two different geographical regions of Turkey in their responses to salinity.
MATERIALS AND METHODS
This experiment was conducted under laboratory conditions of Horticulture Department, Agriculture Faculty, Namik Kemal University in Turkey in 2005.
Plant materials: In this research, it was utilized from two ecotypes of V. vinifera subsp. sylvestris (C.C. Gmelin) which are grows in Marmara Region and Akdeniz Region of Turkey. Meanwhile, 5 BB Kober (V. berlandieri Planch. x V. riparia Michx.) which generally well adapts to most of viticulture regions of Turkey and Isabella grape (V. labrusca L.) which naturally grows on its own root system in East Black Sea Coast of Turkey, were also added to this experiment to make a comparison.
Salinity treatments, analyses, calculations, measurements and tests: Dormancy is a phenomenon occurring in seeds and can be also create problems in most of species’ seed germinations. For this reason, extracted seeds from grape berries were stratified to be permeable to water by humidified sand to break dormancy for 3 months (+4 °C) (Celik, 1998). Prior to germination, seeds were initially surface-sterilized with 2% sodium hypoclorite for 15 min and well washed out with running tap water and demineralized water, respectively. In order to determine salinity tolerance of V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes, 5 BB Kober and Isabella grape, NaCl was added at 0, 2.7, 5.4, 8.1 and 10.8 dS m-1 to nutrient solution to obtain five salinity levels (Aydemir and Ince, 1988). Afterwards, seeds were germinated in rolls of neutral pH germ test paper partially immersed in 1/5 strength Clark’s Nutrient Solution, having 5.5 pH value (Feitosa de Lacerda et al., 2003). 10 seeds for representation of each region were put in rolls of neutral pH germ test paper and rolls with artificially salinized Clark’s Nutrient Solution were put in germination cabinet for germination test and kept at 20/30°C (16/8 h) (Anonymous, 2005) throughout 28 day (Riley, 1981). At the end of germination phases, germination percentages were calculated and no germination was observed under the stress conditions induced by 10.8 dS m-1 NaCl treatment during the germinations of all seeds. Later, plantlets were taken out and roots and shoots (along with cotyledons) were separated from the seeds and fresh weight (FW) and dry weight (DW) of shoots and roots were determined. In order to determine dry weights, shoots and roots were dried at 55°C for 48 hours in an oven. Water content (WC) was calculated using following formula according to Misra and Dwivedi (2004).
Water Content (WC) = ((FW-DW)/FW)x100 |
Moreover, a tolerance index (TI) was calculated for V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes, 5 BB Kober, Isabella grape using the following formula adopted by Maiti et al. (1996).
The oven dried samples were ground to fine powder and 15 mg of this transferred to a digestion flask (25 mL) containing an acid mixture of HNO3 and HClO4, in the ratio
2:1 (v/v). The flask was heated gently over a sand bath. The cooled digest was then diluted by adding distilled water and the volume was made up as required. The contents of K+ and Na+ were found out using flame photometer following the methods described by Tekeli et al. (2003). Each treatment was analyzed with 3 replicates and standard deviation (SD) was calculated (Korkut, 1992).
Statistical analysis: Experiment was set up according to completely randomized block design with 3 replicates of each treatment including 10 seeds per replicate (Korkut, 1992).
Statistical analysis were made using the TARIST statistical program (Anonymous, 1982) and differences among the averages in related criteria were determined using LSD test at 5% significant level.
RESULTS
Germination percentages of seeds showed variations under the different salinity stress. During the germination, first radicles appeared on 7th day of germination phase for Marmara Region, Akdeniz Region, 5 BB Kober and on 16th day of germination phase for Isabella grape depending on NaCl levels (Fig. 1-4). As regards germination phases, it was seen that percentages remained to be low such as 20% for Marmara Region, 20% for Akdeniz Region, 13% for Isabella grape, 10% for 5 BB Kober in all control treatments than NaCl treatments. As shown in Fig. 1-4, however, germination percentages of seeds under the conditions of salinity stress, increased along with rising salinity levels at 33.30% for 2.7 dS m-1, 100% for 5.4 dS m-1, 93.30% for 8.1 dS m-1 in Marmara Region; 10% for 2.7 dS m-1, 30% 5.4 dS m-1, 93.30% for 8.1 dS m-1 in Akdeniz Region; 10% for 2.7 dS m-1, 6.70% for 5.4 dS m-1, 20% for 8.1 dS m-1 in Isabella grape and 26.70% for 2.7 dS m-1, 93.30% for 5.4 dS m-1, 3.30% for 8.1 dS m-1 in 5 BB Kober, respectively. Towards the end of germination phases in seeds, the highest germination percentages were obtained from 5.4 and 8.1 dS m-1 in Marmara Region (100% and 93.30%); 8.1 dS m-1 in Akdeniz Region (93.30%); 5.4 dS m-1 in 5 BB Kober (93.30%). On the other hand, the highest germination percentage in Isabella grape was at 20% for 8.1 dS m-1.
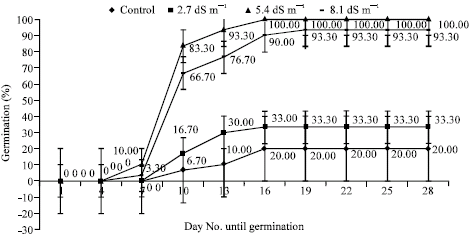 | |
| Fig. 1: | Effects of salinity on germination percentages of Marmara Region seeds. Each value shows average of 3 independent observations and standard deviation was determined |
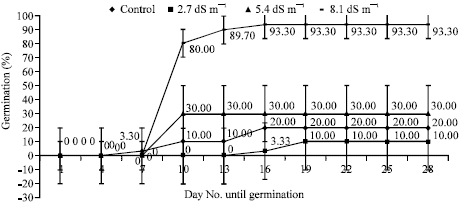 | |
| Fig. 2: | Effects of salinity on germination percentages of Akdeniz Region seeds. Each value shows average of 3 independent observations and standard deviation was determined |
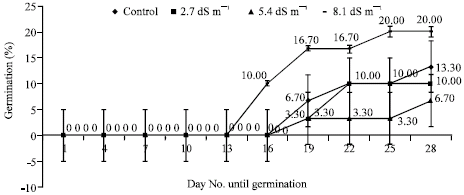 | |
| Fig. 3: | Effects of salinity on germination percentages of Isabella grape seeds. Each value shows average of 3 independent observations and standard deviation was determined |
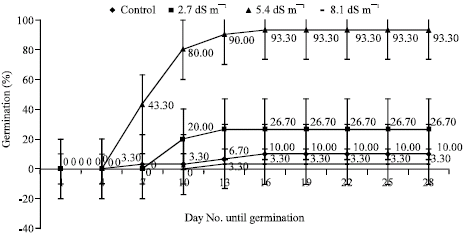 | |
| Fig. 4: | Effects of salinity on seed germination percentages of 5 BB Kober seeds. Each value shows average of 3 independent observations and standard deviation was determined |
At the end of germination phase, some analyses, calculations, measurements were performed on shoots and roots of plantlets emerged from the seeds. Regarding averages of shoot fresh weight in plantlets, the highest values were 110.30 mg for control in Marmara Region, 79.50 mg for control in Akdeniz Region, 84.50 mg for 2.7 dS m-1 in Isabella grape, 87.30 mg for control in 5 BB Kober. On the other hand, the highest values of root fresh weight were 16.40 mg for 2.7 dS m-1 and 5.4 dS m-1 in Marmara Region, 15.70 mg for 2.7 dS m-1 in Akdeniz Region, 13.40 mg for control in Isabella grape, 40.80 mg for 2.7 dS m-1 in 5 BB Kober (Table 1).
As pointed out in Table 1, while the highest values of shoot dry weight were 7.30 mg for 2.7 dS m-1 in Marmara Region, 6.00 mg for 2.7 dS m-1 in Akdeniz Region, 10.20 mg for 2.7 dS m-1 in Isabella grape, 6.20 mg for 8.1 dS m-1 in 5 BB Kober; the highest values of root dry weight became 2.10 mg for 8.1 dS m-1 in Marmara Region, 1.90 mg for 2.7 dS m-1 in Akdeniz Region, 2.20 mg for control in Isabella grape, 1.30 mg for control in 5 BB Kober.
With respect to water content of shoot, the highest values were 94.29% for control in Marmara Region, 94.21% for control in Akdeniz Region, 88.73% for 8.1 dS m-1 in Isabella grape, 94.50% for control in 5 BB Kober. Meanwhile, the highest water content values of root were 93.29% for 5.4 dS m-1 in Marmara Region, 88.46% for control in Akdeniz Region, 90.15% for 8.1 dS m-1 in Isabella grape, 97.30% for 2.7 dS m-1 in 5 BB Kober (Table 1).
As indicated in Table 1, Na+:K+ values of plantlet shoots and roots were lower in control treatments and rose with increasing salinity levels in shoots and roots of plantlets. The highest Na+:K+ values of shoots were 0.53 for 8.1 dS m-1 in Marmara Region, 0.56 for 8.1 dS m-1 in Akdeniz Region, 0.48 for 8.1 dS m-1 in Isabella grape, 0.58 for 8.1 dS m-1 in 5 BB Kober. Besides, the highest Na+:K+ values of roots were 0.72 for 8.1 dS m-1 in Marmara Region, 0.71 for 8.1 dS m-1 in Akdeniz Region, 0.69 for 8.1 dS m-1 in Isabella grape, 0.71 for 8.1 dS m-1 in 5 BB Kober.
As far as tolerance index of shoot length in plantlets was concerned, variations were observed depending on NaCl levels (Fig. 5). Average values were 1.04 in Marmara Region, 0.78 in Akdeniz Region, 0.40 in Isabella grape, 0.77 in 5 BB Kober for 2.7 dS m-1; 0.33 in Marmara Region, 0.62 in Akdeniz Region, 0 in Isabella grape, 0.35 in 5 BB Kober for 5.4 dS m-1; 0.33 in Marmara Region, 0.23 in Akdeniz Region, 0.39 in Isabella grape, 0.25 in 5 BB Kober for 8.1 dS m-1.
| Table 1: | Effects of salinity on fresh weight (mg), dry weight (mg), water content (%) and Na+:K+ values in shoots and roots of plantlets derived from V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes, Isabella grape and 5 BB Kober |
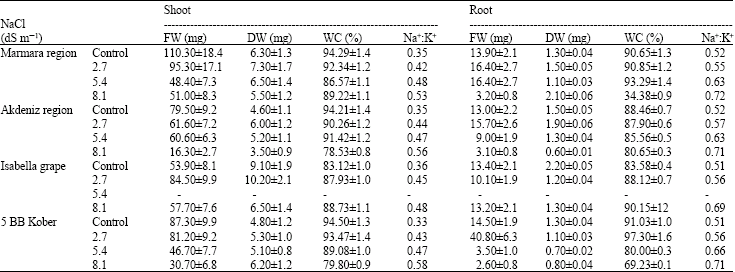 | |
| *Each value shows average of 3 replicates and they were rounded up to nearest figure and standard deviation was determined; FW: Fresh Weight; DW: Dry Weight; WC: Water Content | |
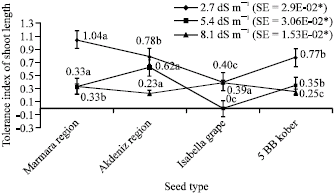 | |
| Fig. 5: | Tolerance index of shoot length depending on different salt treatments (*p<0.05) |
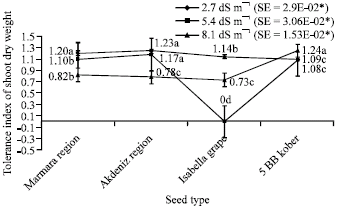 | |
| Fig. 6: | Tolerance index of shoot dry weight depending on different salt treatments (*p<0.05) |
As shown in Fig. 6, tolerance index values of shoot dry weight in plantlets were respectively 1.20 in Marmara Region, 1.23 in Akdeniz Region, 1.14 in Isabella grape, 1.09 in 5 BB Kober for 2.7 dS m-1; 1.10 in Marmara Region, 1.17 in Akdeniz Region, 0 Isabella grape, 1.08 in 5 BB Kober for 5.4 dS m-1; 0.82 in Marmara Region, 0.78 in Akdeniz Region, 0.73 in Isabella grape, 1.24 in 5 BB Kober for 8.1 dS m-1.
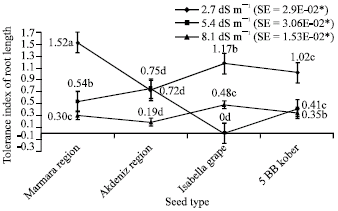 | |
| Fig. 7: | Tolerance index of root length depending on different salt treatments (*p<0.05) |
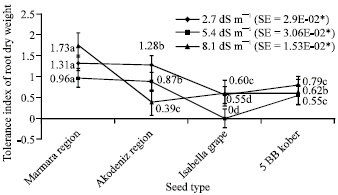 | |
| Fig. 8: | Tolerance index of root dry weight depending on different salt treatments (*p<0.05) |
Results about tolerance index of root length in plantlets were presented Fig. 7. From this standpoint, average values were 1.52 in Marmara Region, 0.72 in Akdeniz, 1.17 in Isabella grape, 1.02 in 5 BB Kober for 2.7 dS m-1; 0.54 in Marmara Region, 0.75 in Akdeniz Region, 0 in Isabella, 0.41 in 5 BB Kober for 5.4 dS m-1; 0.30 in Marmara Region, 0.19 in Akdeniz Region, 0.48 in Isabella grape, 0.35 in 5 BB Kober for 8.1 dS m-1.
Figure 8 shows tolerance index of root dry weight in plantlets. Depending on NaCl, averages were respectively 1.31 in Marmara Region, 1.28 in Akdeniz Region, 0.55 in Isabella grape, 0.79 in 5 BB Kober for 2.7 dS m-1; 0.96 in Marmara Region, 0.87 in Akdeniz Region, 0 in Isabella grape, 0.55 in 5 BB Kober for 5.4 dS m-1; 1.73 in Marmara Region, 0.39 in Akdeniz Region, 0.60 in Isabella grape, 0.62 in 5 BB Kober for 8.1 dS m-1 (Fig. 8).
DISCUSSION
V. vinifera L. is well known to be resistant to soil salinity than most of grapevine rootstocks and V. vinifera subsp. sylvestris (C.C. Gmelin) is a subspecies of V. vinifera L. Salinity studies about viticulture showed that although salinity levels of vineyard soil between 2.03 and 3.38 dS m-1 had non-significant effects on crop losses of grapevine (less than 0.10), other salinity levels between 3.38 and 5.40 dS m-1 resulted in significant crop losses (between 0.10 and 0.25) (Anonymous, 1978).
Ungar (1982 and 1991) stated that salinity reduced the total number of germinated seeds and delayed initiation of germination processes; the responses to salinity were variable and species specific within each group. Ellis et al. (1983) declared that seed germination percentage of Vitis ssp. remained to be low. Salinity and temperature interact in their control of seed germination (Khan and Ungar, 1999), with the greatest inhibition due to salinity usually found at the minimum or maximum limits of tolerance to temperature (Badger and Ungar, 1989). The dependence of the response to salinity on temperature may be due to an initial inhibition of germination at higher salt concentrations at some temperatures rather than others (De Villiers et al., 1994; Khan and Ungar, 1997, 1998, 1999; Khan and Gul, 1999), while alternating day and night temperatures have been shown to promote germination in a number of halophytes (Okusanya, 1977; Ungar, 1995; Khan and Ungar, 1997, 1998, 1999; Gul and Weber, 1999).
In this study, germination percentages in all control treatments were remained to be low and gradually risen germination percentages were obtained from NaCl treated seeds at moderate temperatures (20/30°C). During the whole germination phase, Isabella grape exhibited low germination percentages because it had hard seed coat and huge seed size.
Parida et al. (2004) informed that plant height, fresh weight and dry matter of plant could not be affected via different salinity levels. As expected, the highest values of shoot and root fresh weights were obtained from control treatments and 2.7 dS m-1 NaCl treatments in our study.
When shoot and root dry weight were examined, it was seen that the highest values were mostly obtained from 2.7 dS m-1 NaCl and control treatments.
Khan (2001) and Romeroaranda et al. (2001) declared that water content in shoot and root was not affected in salinity conditions. As far as they are concerned, water content and osmotic potential of plants become inversely proportional with an increase in salinity despite the fact that turgor pressure increases with increasing salinity. Parida et al. (2003) informed that water content was reduced by salinity stress conditions in contrast to the leaves of salt treated plants accumulated more water than the leaves of control plants explained by Gebauer et al. (2004). Our research results about water contents in shoot and root of plantlets were consistent with Parida et al. (2003). While the highest values of water content in shoot were mostly obtained from control treatments in shoots, values of water content in root slightly showed increase in NaCl treated plantlets than control treatments.
Alian et al. (2000) reported that Na+:K+ ratio could offer a chance to predict crop tolerance for stress as the increase of Na+ in salt tolerant species was generally associated with reduce in K+. From Na+:K+ averages of shoot and root standpoint, it was observed that average values increased with rising NaCl levels. As far as Na+:K+ values of roots were concerned, it was seen that the most salinity tolerant plantlets respectively belonged to Marmara Region, Akdeniz Region and 5 BB Kober, Isabella grape for 8.1 dS m-1 NaCl treatment.
Plantlets originated from Marmara Region, Akdeniz Region, Isabella grape and 5 BB Kober exhibited diversity in tolerance indexes under salinity stress conditions. Plantlets showing a high tolerance index for shoot length under 8.1 dS m-1 NaCl treatment were respectively Isabella grape, Marmara Region, 5 BB Kober, Akdeniz Region.
Plantlets derived from Isabella grape, 5 BB Kober, Marmara Region, Akdeniz Region showed high tolerance indexes for root length under 8.1 dS m-1 NaCl treatment, respectively.
5 BB Kober plantlets had the highest tolerance index for shoot dry weight under 8.1 dS m-1 NaCl treatment and Marmara Region, Akdeniz Region and Isabella grape followed this sequence.
As for tolerance index of root dry weight, Marmara Region, 5 BB Kober, Isabella grape and Akdeniz Region represented high tolerance index under 8.1 dS m-1 NaCl treatment, respectively. Diversities were found in all plantlet types and even both V. vinifera subsp. sylvestris (C.C. Gmelin) ecotypes in salinity tolerances and plantlets originated from Marmara Region were found to be salinity resistant than Akdeniz Region.
All in all, results have shown that V. vinifera subsp. sylvestris (C.C. Gmelin) seeds from Marmara Region should be used to obtain salinity resistant hybridized grape rootstocks in breeding programs of viticulture and rooted plant materials of V. vinifera subsp. sylvestris (C.C. Gmelin) obtained from Marmara Region by means of vegetative propagation methods as grapevine rootstock can be also preferred by grape growers for salinity soils.
REFERENCES
- Alian, A., A. Altman and B. Heuer, 2000. Genotypic difference in salinity and water stress tolerance of fresh market tomato cultivars. Plant Sci., 152: 59-65.
CrossRef - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Bohnert, H.J., D.E. Nelson and R.G. Jensen, 1995. Adaptations to environmental stresses. Plant Cell., 7: 1099-1111.
CrossRefPubMedDirect Link - Gebauer, J., K. El-Siddig, A.A. Salih and G. Ebert, 2004. Tamarindus indica L. plantlets are moderately salt tolerant when exposed to NaCl induced Salinity. Scientia Horticulturae, 103: 1-8.
CrossRefDirect Link - Gul, B. and D.J. Weber, 1999. Effect of salinity, light and temperature on germination in Allenrolfea occidentalis. Can. J. Bot., 77: 240-246.
Direct Link - Khan, M.A. and I.A. Ungar, 1984. Seed polymorphism and germination responses to salinity stress in Atriplex triangularis willd. Bot. Gaz., 145: 487-494.
Direct Link - Khan, M.A. and I.A. Ungar, 1997. Effect of thermoperiod on recovery of seed germination of halophytes from saline conditions. Am. J. Bot., 84: 279-283.
Direct Link - Khan, M.A. and I.A. Ungar, 1998. Germination of salt tolerant shrub Suaeda fruticosa from Pakistan: Salinity and temperature responses. Seed Sci. Technol., 26: 657-667.
Direct Link - Aziz, I. and M.A. Khan, 2001. Experimental assessment of salinity tolerance of Ceriops tagal seedlings and saplings from the Indus delta, Pakistan. Aquatic Bot., 70: 259-268.
CrossRef - Lin, C.C. and C.H. Kao, 1995. NaCl stress in rice seedlings: The influence calcium on root growth. Bot. Bull. Acad. Sin., 36: 41-45.
Direct Link - Maiti, R.K., L.E.D. Amaya, S.I. Cardona, A.M.O. Dimas, M. De La Rosa-Ibarra and H.D. Castillo, 1996. Genotypic variability in maize cultivars (Zea mays L.) for resistance to drought and salinity at the seedling stage. J. Plant Physiol., 148: 741-744.
Direct Link - Misra, N. and U.N. Dwivedi, 2004. Genotypic difference in salinity tolerance of green gram cultivars. Plant Sci., 166: 1135-1142.
CrossRefDirect Link - Parida, A.K., A.B. Das and B. Mittra, 2004. Effects of salt on growth, ion accumulation, photosynthesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees, 18: 167-174.
CrossRefDirect Link - Parida, A.K. and A.B. Das, 2005. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf., 60: 324-349.
CrossRefPubMedDirect Link - Ribaut, J.M., C. Jiang, D. Gonzalez-de-Leon, G.O. Edmeades and D.A. Hosington, 1997. Identification of quantitative trait loci under drought conditions in tropical maize, 2. Yield components and markerassisted selection strategies. Theor. Applied Genet., 94: 887-896.
CrossRefDirect Link - Romeroaranda, R., T. Soria and J. Cuartero, 2001. Tomato plant water uptake and plant water relationship under saline growth conditions. Plant Sci., 160: 265-272.
Direct Link - Qadir, M., N. Qureshi, N. Ahmad and N. Ahmad, 1997. Nutrient availability in a calcareous saline-sodic soil during vegetative bioremediation. Arid Soil Res. Rehab., 11: 343-352.
CrossRefDirect Link - Sreenivasulu, N., S.K. Sopory and P.B.K. Kishor, 2007. Deciphering the regulatory mechanisms of abiotic stress tolerance in plants by genomic approaches. Gene, 388: 1-13.
CrossRefDirect Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link