
M.A. Hassanain
Department of Zoonotic Diseases
Mona S. Mahmoud
Department of Parasitology and Animal Diseases, Veterinary Research Division, National Research Center, Post Box 12622, El-Tahrir Street Dokki, Giza, Egypt
Nawal A. Hassanain
Department of Zoonotic Diseases
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 13 | Page No.: 2123-2129







ABSTRACT
Serum samples collected from a total number of twenty parasitologically confirmed cases of human fasciolosis were used to evaluate the diagnostic sensitivity and specificity of snail derived antigens; non infected snail (SA), infected snail (ISA), redia (RA), cercaria (CA) and encysted metacercaria (EMCA) of F. gigantica using the enzyme linked immunosorbent assay (ELISA) and enzyme linked immunotransfer blot (EITB). ELISA results showed that the highest level of sensitivity (100%) was with the CA as compared with ISA, RA and EMCA which displayed lower sensitivity levels of 30, 60 and 90%, respectively. All fasciolosis patients were seronegative with SA. Sodium dodecyle sulfate polyacrylamide gel electrophoresis and EITB (with rabbit antiserum raised against F. gigantica somatic antigen) of the snail derived antigens were carried out to characterize their protein profiles and detect cross-reactive polypeptide bands between them. The immunoblot profile of SA displayed cross-reactive bands at 61 and 30 kDa with ISA, RA, CA and EMCA; CA at 61, 34 and 30 kDa with ISA; 96, 61, 34, 30 and 23 kDa with RA and 61, 34, 23 and 19 kDa with EMCA. Cross reactive antigens may be important as possible candidates for vaccine and diagnosis of fasciolosis. Immunoblot of sera from fasciolosis patients using fractionated cercarial antigen on nitrocellulose strips showed that all sera recognized common reactive band at 32.5 kDa molecular weight from cercarial antigen. We suggest that the 32.5 kDa component of Fasciola cercarial antigen may be the most sensitive and specific for the diagnosis of human fasciolosis.
PDF Abstract XML References Citation
How to cite this article
M.A. Hassanain, Mona S. Mahmoud and Nawal A. Hassanain, 2007. Evaluation of Snail Derived Antigens of Fasciola gigantica for Serodiagnosis of Human Fasciolosis. Pakistan Journal of Biological Sciences, 10: 2123-2129.
DOI: 10.3923/pjbs.2007.2123.2129
URL: https://scialert.net/abstract/?doi=pjbs.2007.2123.2129
DOI: 10.3923/pjbs.2007.2123.2129
URL: https://scialert.net/abstract/?doi=pjbs.2007.2123.2129
INTRODUCTION
Fasciolosis caused by Fasciola hepatica and Fasciola gigantica is one of the major public health problems in the world. Fasciolosis was added to the list of important helminthiases with a great impact on human development at the third Global meeting of the partners for parasite control held in WHO Head quarter Geneva in November 2004 (Mas-Coma et al., 2005).
Fasciolosis is not only an important human disease but also affect many herbivorous mammals. Animal fasciolosis causes world wide economic losses of approximately two billion dollars per year (Torgerson and Claxton, 1999). Tropical fasciolosis is the most economically important helminthic infection of ruminant in Asia and Africa (Spithill et al., 1997).
Human and animal fasciolosis is an increasing worldwide zoonotic liver fluke disease. In general, human fasciolosis was very sporadic until the last three decades where clinical cases and outbreaks were reported. The present climate and global change appear to increasingly affected snail born helminthes, which are strongly dependent on environmental factors for dissemination. The total estimated number of infected people is 2.4 million in 61 countries and the number at risk is more than 180 million throughout the world (Mas-Coma et al., 1999). Different diagnostic limitations and the fact that human fasciolosis is not a disease of obligatory declaration suggest that the number of human cases is much greater than published (Mas-Coma et al., 2005).
Fasciolosis is one of the most important infections in Egypt. There are currently 830,000 people suffering from fasciolosis (Haseeb et al., 2002). Human infection is one of the main causes of hepatic disorders in Egypt (Malked et al., 1988). Man is infected through the ingestion of encysted metacercariae attached to vegetables (Pantelouris, 1965). Adult worms of Fasciola spp. live in the bile ducts of the liver and may lead to liver cirrhosis.
Clinically, human fasciolosis may be false diagnosed as amoebic hepatitis, acute infective hepatitis as well as disorders associated with cirrhosis and primary carcinoma human fasciolosis based on the demonstration of the eggs in stool, duodenal contents or bile is usually unsatisfactory due to false passage of eggs, ectopic fasciolosis and failure of immature worms to maturation. In addition, the use of previously reported diagnostic assays for fasciolosis demonstrated the presence of strong cross-reactivity between Schistosoma mansoni and Fasciola hepatica infections (Hillyer, 1979, 1981; Christensen et al., 1978, 1980).
Various techniques have been evaluated for diagnosis of human fasciolosis using different Fasciola antigenic preparations, including partially purified antigen and excretory-secretory product of adult worms (El-Kerdany et al., 2002; Intapan et al., 2003; Tantrawatpan et al., 2003; Dalimi et al., 2004). However, these serological assays still lack the diagnostic specificity, especially in areas where more than one helminthic infection prevails. This lack of specificity in the serodiagnosis of fasciolosis may be due to the use of antigens obtained from adult worm stages.
Little is known of the identity and characterization of Fasciola specific antigens within their intermediate snail host (Weston et al., 1994; El Bahy and Shalaby, 2004).
The present study aimed to evaluate snail derived antigens; non infected snail (SA), infected snail (ISA), redia (RA), cercaria (CA) and encysted metacercaria (EMCA) using two sensitive tests; enzyme linked immunosorbent assay (ELISA) and enzyme linked immunotransfer blot (EITB) to detect the most sensitive and specific antigen for diagnosis of human fasciolosis.
MATERIALS AND METHODS
Subjects: A total of twenty fasciolosis patients and eight controls (parasite-free individuals) were selected. To ensure infection with Fasciola in fasciolosis patients, stool samples were examined by Kato thick smear (Katz et al., 1970) and or fluke finder technique (Welch et al., 1987). Rectal snip was done for fasciolosis patients and controls to ensure the absence of schistomiasis in both groups. Medical sheets were filled on each patient and control. Serum samples were collected from both fasciolosis patients and controls and stored at -20°C till used.
Parasitological studies
Collection of cercariae: F. gigantica eggs were collected from the gall bladders of naturally infected buffaloes or cattle in Cairo abattoir. After several washing with saline, the eggs were incubated at 28°C in a dark place for 13 days following the protocol of Bory (1964). After egg hatching in direct sun light, the shedded miracidiae were collected. The collected miracidiae were used to infect Lymnaea cailliaudi snails which were reared and grown in the laboratory according to the method described by Augot et al. (1997) to obtain cercariae.
Collection of metacercariae: The recovered cercariae were collected in Petri-dishes lined with cellophane containing dechlorinated tap water for encystment at room temperature (Rajase-Kariah et al., 1979). Microscopically, the viability of the metacercariae was tested based on the movements of the young flukes inside the cyst and presence of excretory granules in bunches. The collected encysted metacercariae were stored at 4°C till used.
Preparation of antigens
Snail derived antigens
Snail antigen: The snails (L. cailliaudi) antigens were prepared according to Khalil et al. (1985). Hepatopancreases of laboratory bred mature non-infected and infected L. cailliaudi snails were obtained and homogenized in equal amount of saline and then centrifuged at 5000 rpm at 4°C for 1 h. The supernatant was concentrated by dialysis bags and used as snail antigens.
Redial, cercarial and encysted metacercarial antigens: Redial, cercarial and encysted metacercarial antigens were prepared according to Hamburger et al. (1989). The collected rediae, cercariae and encysted metacercariae were sonicated in Phosphate Buffer Saline (PBS) pH 7.2. Sonication was carried out with a sonicator under 225 watts in 3 cycles of pulses for 3-5 min each. The sonicated antigen were subjected to high speed centrifugation (10,000 rpm) for 1 h at 4°C. The supernatant was separated as antigen. Protein content of each prepared antigen was determined by the method of Lowry et al. (1951). Each antigen was aliquoted and stored at -20°C until used.
Fasciola gigantica somatic antigen: Whole worm extracts from adult F. gigantica flukes were prepared as described by Oldham (1983). After 4-5 times washing of the live adult flukes in physiological saline, the flukes were homogenized (2 flukes/mL) using a tissue homogenizer. The homogenate was then subjected to ultrasonication at maximum amplitude (peak to peak) for 10 min, two times, with an interval of 1 min using an Ultrasonicator (Misonix, USA). After sonication, the preparation was centrifuged at 12,000 rpm for 45 min at 4°C. The supernatant was collected and designated as somatic antigen of F. gigantica (FgA) and its protein concentration was measured as described by Lowry et al. (1951).
Preparation of rabbit anti-sera against Fasciola somatic antigen: Two white New-Zealand rabbits were immunized subcutaneously (0.05 mg protein antigen/rabbit) with crude extract of FgA emulsified in equal volume of Freund's complete adjuvant and two rabbits were kept as control. Two weeks later, three booster injections in Freund's incomplete adjuvant were given with one week interval (Alkarmi and Faubert, 1985). Serum samples were collected 4 days after the last booster injection. The serum was designated RaFgA.
Analysis of antigens and antibodies
Enzyme linked immunosorbent assay (ELISA): ELISA was carried out according to Zimmerman et al. (1982). The optimal antigen concentration, antibody and conjugate dilutions were chosen after preliminary checker board titration. In the present study, the optimum conditions were 10 μg mL-1 coating buffer antigen concentration, 1:100 serum dilutions and 1:250 alkaline phosphatase labeled rabbit anti-human IgG (Sigma Co.) as conjugate and 1 mg p-nitrophenyl phosphatase dissolved in 1 mL substrate buffer as substrate. The absorbance of the colored reaction was read within 30 min at 405 nm using a titertek multiskan ELISA reader. All incubation steps were carried out at 37°C in a moist chamber. The positive threshold value was determined to be two-fold the mean cut-off value of negative sera.
Sodium dodecyle sulfate polyacrylamide gel electrophoresis (SDS-PAGE): The gel cast comprised 12% resolving and 4% stacking gels with applied 10 μg/well of the different antigens; SA, ISA, RA, CA and EMCA. Mini-protein II Dual slab cell (Bio-Rad Labs, Richmond, CA) was used to conduct electrophoresis using discontinuous system of Laemmli (1970). The fractionated antigens were visualized by Commassie staining. Analysis of the separated bands was performed by soft ware analysis (Gel Proanalyser).
Enzyme linked immunotransfer blot (EITB): The fractionated snail derived antigens were electrically transferred onto Nitrocellulose (NC) membrane. NC sheets were cut into 0.5 cm strips (Towben et al., 1979) followed by blocking in 5% BSA in PBS for 2 h on a rocker platform. Sera diluted at 1:100 in 5% BSA/PBS-T were reacted with fractionated snail derived antigens NC strips for 2 h on a rocker platform. Following washing, alkaline phosphatase-labeled anti-rabbit IgG and alkaline phosphatase-labeled anti-human IgG (Sigma Co.) diluted at 1:1000 in PBS-T were added to snail derived antigens and cercarial antigen alone NC strips, respectively for 1 h on a rocker platform. The chromogen BCIP/NBT substrate was added to NC strips and allowed to develop for 30 min. The reaction was visualized by the naked eye.
RESULTS
The diagnostic sensitivity of different snail derived antigens of Fasciola gigantica was evaluated by ELISA using sera from patients with parasitologically confirmed fasciolosis (n = 20). Cercarial antigen showed the highest degree of sensitivity, 100%. Infected snail, redial and encysted metacercarial antigens displayed lower sensitivity levels of 30, 60 and 90%, respectively (Fig. 1). All fasciolosis patients (n = 20) were seronegative with snail derived antigen (SA). Cut off O.D value was 0.3 for all the snail derived antigens.
Commassie staining of the electrophoretically migrating proteins from SA, ISA, RA, CA and EMCA revealed approximately 5 to 11 bands ranging from 101- 14.3 kDa (Fig. 2). Structural homology between SA, ISA and RA antigens was apparent especially in term of 89, 61 and 23 kDa and between SA and CA was in term of 53, 34 and 28 kDa .
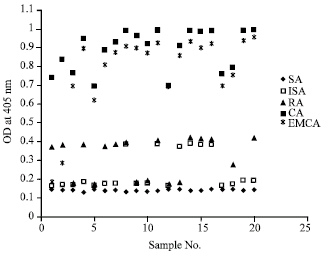 | |
| Fig. 1: | OD values of ELISA Fasciola snail derived antigens of fasciolosis patients (cut off value: 0.3) |
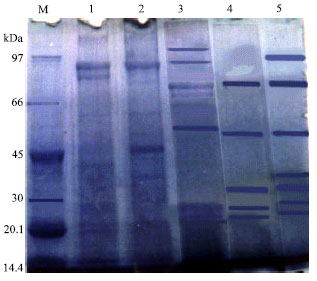 | |
| Fig. 2: | SDS-PAGE pattern of snail derived antigens. M: molecular weight standard, 1: SA, 2: ISA, 3: RA, 4: CA, 5: EMCA |
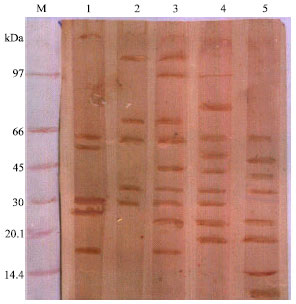 | |
| Fig. 3: | Identification of cross-reactive polypeptide bands recognized by rabbit anti-FgA using EITB. M: molecular weight standard, 1: SA; 2: ISA; 3: RA; 4: CA; 5: EMCA |
 | |
| Fig. 4: | Western blot showing reactivity of Fasciola gigantica cercarial antigen with sera of fasciolosis patients. M: molecular weight standard, 1, 2, 3 and 4: sera of fasciolosis patients |
Two components of 53 and 34 kDa were found in SA and EMCA, while bands of 41 and 17 kDs were only observed in SA and ISA, respectively.
The anti serum raised in rabbits against FgA extract (Ra FgA) was utilized in EITB to identify the specific and the cross reacting antigens (SA, ISA, RA, CA and EMCA) (Fig. 3). Evaluation of the recognized bands from SA revealed that RaFgA recognized 6 polypeptides of M.W. 125, 62, 54, 30, 25 and 17 kDa. Five components of M.W 108, 70, 61, 34 and 29 kDa were identified in ISA. Nine immunoreactive bands (108, 96, 70, 61, 44, 34, 29, 23 and 17 kDa) were detected in RA. Also, nine bands of M.W 96, 77, 61, 50, 42, 34, 29, 23 and 19 kDa were identified in CA. EMC antigen displayed 8 polypeptides of 61, 47, 40, 34, 23, 19, 14 and 12 kDa.
Immunoblot analysis was performed using sera from patients with fasciolosis to investigate whether such sera could recognize defined immunogenic band (s) of electrophoresed cercarial antigen (used as it gave the highest sensitivity in ELISA) for diagnostic purposes. All sera obtained from parasitologically confirmed fasciolosis cases recognized an immunodominant band of 32.5 kDa (Fig. 4).
DISCUSSION
This study aimed at getting specific antigen for accurate diagnosis and consequently proper management of fasciolosis which would obviate many of the complications usually attributed to other causes. Diagnosis of fasciolosis during the patent period is confirmed by direct fecal sedimentation which is not always reliable. That is mainly due to the fluctuating egg deposition which thus requires repeated stool examinations. Also, the fecal examinations are unsuitable for diagnosis of fasciolosis during the pre-patent period which is three months (Soulsby, 1982).
In the present study, non infected snails (SA), infected snails (ISA), redial (RA), cercarial (CA) and encysted metacercarial (EMCA) antigens were used in terms of capturing anti-Fasciola antibodies in sera from fasciolosis patients. ELISA results showed that the cercarial antigen demonstrated the highest degree of sensitivity (100%) as compared to SA (0%), ISA (30%), RA (60%) and EMCA (90%). The decreased sensitivity observed when the different parasite stages were evaluated by ELISA suggested that anti-Fasciola antibodies detected in sera from patients with fasciolosis were mainly against cercarial antigen.
In the current study, structural homology between non infected snails, infected snails, redial, cercarial and encysted metacercarial antigens was apparent by the use of SDS-PAGE. This homology resided in components of similar molecular weights between them as 89, 61 and 23 kDa (SA, ISA and RA), 53, 34 and 28 kDa (SA and CA) and 53 and 34 kDa (SA and EMCA). El-Bahy and Shalaby (2004) observed structural homology that resided in components of similar molecular weights (85, 54, 45, 34, 28 and 22 kDa) between L. cailliaudi snail and F. gigantica miracidial antigens.
Cross-reaction between different stages in the life cycle of F. gigantica with extracts of snail antigens from its intermediate host snail L. cailliaudi was proved in the present study by the use of EITB in which anti sera raised in rabbits against FgA extract (RaFgA) were utilized. Common reactive bands were present in SA and all snail derived antigens (ISA, RA, CA and EMCA) with 61 and 30 kDa. Immunoblot profile of ISA and CA displayed common reactive bands at 61, 34 and 29 kDa; RA and CA at 96, 61, 34, 29 and 23 kDa and EMCA and CA at 61, 34, 23 and 19 kDa. This high similarity in antigenic composition between the snail and the species of trematode stages able to develop inside it is in agreement with the finding of Henning et al. (1978) that immuno electrophoresis has showed cross reaction between different stages in the life cycle of S. mansoni and extracts from intermediate host snails B. glabrata and B. alexandrina. EL-Bahy and Shalaby (2004) observed intensive cross reaction between L. cailliaudi snail and F. gigantica miracidium. They also recorded less cross reactivity with incompatible miracidium by use of immunoblot in which rabbit anti miracidium antisera were utilized.
Santiago de Weil and Hillyer (1986) reported the use of immunodominant molecular weight bands of 31 and 33 kDa (adult worm antigen) in the diagnosis of fasciolosis. Also, studies performed using the E/S products from the adult worm have revealed immunogenic polypeptide bands (25 and 27 kDa) derived from F. hepatica (Santiago de Weil and Hillyer, 1988; Yamasaki et al., 1989; Sampaio-Silva et al., 1996) and from F. gigantica (27 kDa) (Mousa, 1992) and (26- to 28-kDa) (Attallah et al., 2002). These bands demonstrated high degrees of specificity when evaluated in the serodiagnosis of fasciolosis. However, these studies did not evaluate the efficacy of antigens derived from the early developmental stages of the parasite.
In the present study, Immunoblot analysis of sera using fractionated cercarial antigen on NC strips, revealed an immunodominant polypeptide band of M.W. 32.5 kDa recognized by all sera from fasciolosis patients. Similarly, previous studies have reported the use of immunodominant molecular weight band of 32.5 kDa (F. gigantica cercarial antigen) in the diagnosis of human (Mousa et al., 1996) and sheep fasciolosis (Mousa, 2001).
CONCLUSIONS
The present study evaluated the use of snail derived antigens of F. gigantica in the serodiagnosis of human fasciolosis. Cercarial polypeptide antigen of 32.5 kDa demonstrated a high level of sensitivity and specificity when tested with sera from patients with fasciolosis and therefore could be considered of diagnostic potential for fasciolosis. Detection of serum antibodies to this defined cercarial antigen as yet, does not discriminate between current and previous infection. Also, we have confirmed the presence of common antigens between F. gigantica stages and its vector, L. cailliaudi. Cross reactive antigens may be important as possible candidates for vaccine and diagnosis of fasciolosis.
REFERENCES
- Attallah, A.M., E.A. Karawia, H. Ismail, A.A. Tabll and A.A. Nawar et al., 2002. Identification and characterization of a 26- to 28-kDa circulating antigen of Fasciola gigantica. Ann. Trop. Med. Parasitol., 96: 271-282.
CrossRefDirect Link - Augot, D., D. Rondelaud, G. Dreyfuss and J. Cabaret, 1997. Fasciola hepatica: In vitro production of daughter rediae and cercariae from first and second generation rediae. Parasitol. Res., 83: 383-385.
Direct Link - Dalimi, A., R. Hadighi and R. Madani, 2004. Partially Purified Fraction (PPF) antigen from adult Fasciola gigantica for the serodiagnosis of human fascioliasis using Dot-ELISA technique. Ann. Saudi Med., 24: 18-20.
PubMed - El-Kerdany, E.D., N.M. Abd-Alla and O.A. Sharaki, 2002. Recognition of antigenic components of Fasciola gigantica and their use in immunodiagnosis. J. Egypt. Soc. Parasitol., 32: 675-690.
Direct Link - Haseeb, A.N., A.M. El-Shazly, M.A. Arafa and A.T. Morsy, 2002. A review of fascioliasis in Egypt. J. Egypt. Soc. Parasitol., 32: 37-54.
Direct Link - Intapan, P.M., W. Maleewong, S. Nateeworanart, C. Wongkham, V. Pipitgool, V. Sukolapong and S. Sangmaneedet, 2003. Immunodiagnosis of human fascioliasis using an antigen of Fasciola gigantica adult worm with molecular mass of 27 kDa by a dot-ELISA. Southeast Asian J. Trop. Med. Public Health, 34: 713-717.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Mas-Coma, S., M.D. Bargues and M.A. Valero, 2005. Fascioliasis and other plant borne trematode zoonoses. Int. J. Parasitol., 35: 1255-1278.
Direct Link - Mousa, W.M., 2001. Evaluation of cercarial antigen for the serodiagnosis of fasciolosis in experimentally and naturally infected sheep. Vet. Parasitol., 97: 47-54.
Direct Link - Oldham, G., 1983. Antibodies to F. hepatica antigen during experimental infection in cattle measured by ELISA. Vet. Parasitol., 13: 151-158.
CrossRef - Santiago, N. and G.V. Hillyer, 1988. Antibody profiles by EITB and ELISA of cattle and sheep infected with Fasciola hepatica. J. Parasitol., 74: 810-818.
CrossRefPubMedDirect Link - Spithill, T.W., D. Piedrafita and P.M. Smooker, 1997. Immunological approaches for the control of fasciolosis. Int. J. Parasitol., 27: 1221-1235.
Direct Link - Tantrawatpan, C., W. Maleewong, C. Wongkham, S. Wongkham, P.M. Intapan and K. Nakashima, 2003. Characterization of Fasciola gigantica adult 27-kDa excretory-secretory antigen in human fascioliasis. Parasitol. Res., 91: 325-327.
Direct Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Welch, R.D., P.H. Smith, J.B. Malone, R.A. Holmes and J.P. Geaghan, 1987. Herd evaluation of Fasciola hepatica infection in Louisiana cattle by an ELISA. Am. J. Vet. Res., 48: 345-347.
PubMed - Weston, D., B. Allen, A. Thakur, P.L. Loverde and W.M. Kemp, 1994. Invertebrate host parasite relationship convergent evaluation of a tropomyosion epitope between Schistosoma sp., Fasciola hepatica and certain pulmonate snails. Exp. Parasitol., 78: 269-278.
Direct Link - Zimmerman, G.L., L.W. Jen, J.E. Cerro, K.L. Farnsworth and R.B. Wescott, 1982. Diagnosis of Fasciola hepatica infections in sheep by an enzyme-linked immunosorbent assay. Am. J. Vet. Res., 43: 207-210.
Direct Link