
Fatemeh Zarinkamar
Faculty of Basic Sciences, Tarbiat Modarres University, Tehran, P. O. Box 14115-175, Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 2 | Page No.: 199-219







ABSTRACT
Since 1998, research into more than 470 plant species from some 65 families, found growing in a unique protected area of Irano-Torany, has examined diverse aspects of their anatomical and structural features and has studied the ecological implications of these finding for each species. The present paper, as a part of this research, focuses on the stomatal characters of 326 species from 36 families of dicotyledons, the majority of which are herbaceous species. The stomatal density, guard cell lengths on the adaxial and abaxial leaf epidermis and the stomatal type in each family is described and the relationship between stomatal density and guard cell size is reviewed. The stomatal characteristics presented here are a valuable research resource, allowing; I) the identification of basic stomatal types in these plants, II) the facilitation of their taxonomic classification and III) constitution of a baseline data set against which to monitor and evaluate environmental changes at these sites of international conservation interest.
PDF Abstract XML References Citation
How to cite this article
Fatemeh Zarinkamar, 2007. Stomatal Observations in Dicotyledons. Pakistan Journal of Biological Sciences, 10: 199-219.
DOI: 10.3923/pjbs.2007.199.219
URL: https://scialert.net/abstract/?doi=pjbs.2007.199.219
DOI: 10.3923/pjbs.2007.199.219
URL: https://scialert.net/abstract/?doi=pjbs.2007.199.219
INTRODUCTION
Stomata are considered to be one of the major structures within the leaf organ that have allowed the higher plants to adapt to virtually all terrestrial environments on the planet, by means of adjustment of their size, density and distribution (Zarinkamar, 2006b). An alteration of leaf stomatal density can be used as an indicator of environmental change (Case, 2004). Several researchers have shown that stomatal densities change in response to water availability (Edward and Meidner, 1978), light intensity (Retallk, 2001; Lu et al., 1993), temperature (Ciha and Brun, 1975), geographical location (Retallk, 2001) and CO2 concentration (Bristow and Looi, 1968; Woodward, 1987; Woodward and Bazzaz, 1988).
As environmental conditions affect stomatal density, this has implications for plant development. Collection of data on stomatal densities from sites which have experienced a minimum of environmental pollution and human impact can provide a baseline against which to assess anthropogenic impacts on these species elsewhere. This has the potential to facilitate appropriate monitoring and analysis of environmental changes, using stomatal characteristic as an indicator of physiological response. Suitable sites for such monitoring could be found amongst protected areas. These areas have experienced minimum levels of local environmental disturbance such as water and air contamination, waste pollution and human intervention.
MATERIALS AND METHODS
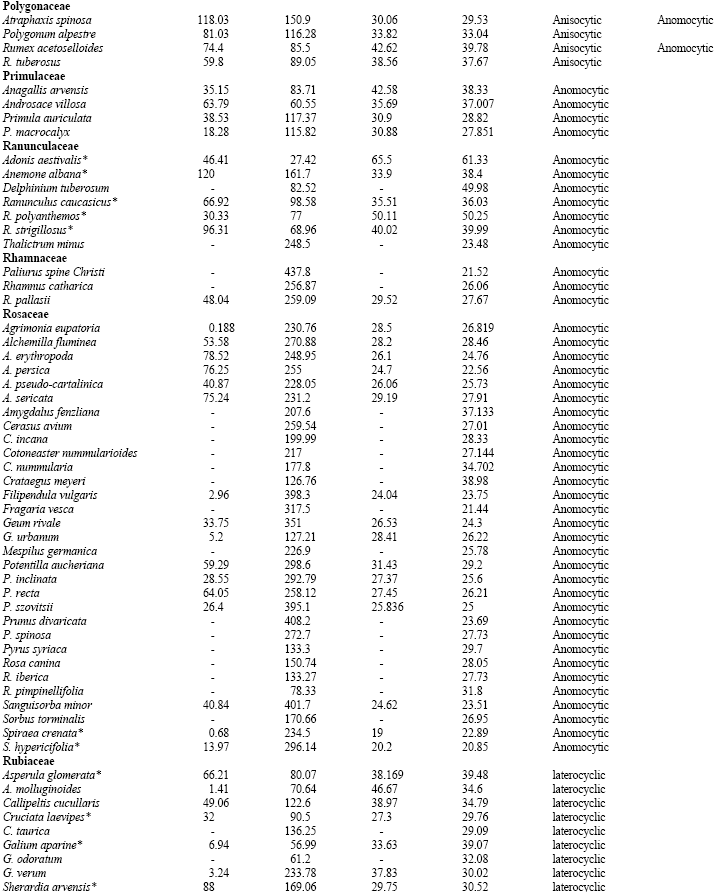
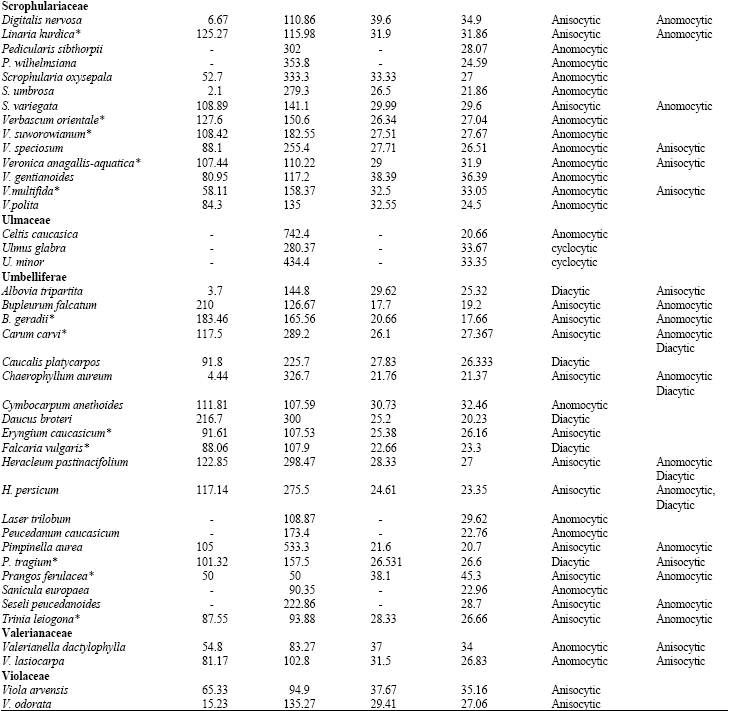
Some 320 species pertaining to 36 families of dicotyledons were selected to represent a broad spectrum of stomatal characteristics. All materials were identified by the Iranian central herbarium (TARI) and Department of Environment (DoE). To minimise misinterpretation, the central area of the leaf lamina from relatively mature leaves was selected for analysis and at each site, the data entry comprises an average of 30 samples taken from 6 plants of the same species. In order to study stomatal density, the diafanization technique was employed Stritmater (1973) and the results were observed using a light microscope. A combination of stomatal classification schemes of Dilcher (1974) and Wilkinson (1979) quoted by Stace (1989), were used as a basis to define the various stomatal forms encountered in this study. Most of stomatal types quoted by Stace are present in the plant families covered in the present report (shown in Fig. 1; A-R).
Most species in the present study are herbaceous; the exceptions are woody plants found in the following families; Aceraceae, Anacardiaceae, Caprifoliaceae, Cornaceae, Corylaceae, Fagaceae, Oleaceae, Rhamnaceae, Rosaceae and Ulmaceae. The study sites also contain the following endemic species: Minuartia lineate, Silene persica (Caryophyllaceae); Centaurea aziziana, C. zuvandica (Compositae); Erysimum subulatum, Sterigmostemum contortuplicatum (Cruciferae), Hypericum linarioides (Hypericaceae); Scrophularia oxysepala (Scrophulariaceae).
 | |
| Fig. 1: | Stomatal types: A1, Anomocytic; A (2,3,4), Staurocytic; B, actinocytic; C, (1,2), cyclocytic; C (3,4), tetracytic; D, amphicyclocytic, E, (1,2,3), anisocytic; E4, amphianisocytic; F, helicocytic; G, (1,2,3), diacytic; H, (1,2), amphianisocytic; I, (1,2), laterocyclic; I3, paracytic; J, (1,2), amphiparacytic; K, bracyparacytic; L, (1,2), amphibracyparacytic; M, hemiparacytic; N, paratetracytic; O, amhiparatetracytic; P, brachuparatetracytic; Q, amphibrachyparatetracytic, R (1,2,3) parahexacytic-monopolar; R4, parahexacytic-dipolar |
RESULTS
Table 1 shows the stomatal densities and characteristics including type of stomata for all species studied. Metcalf and Chalk (1950) listed type of stomata in dicotyledons and based on that, Table 1 develops dominate and subordinate type of stomata as well as checking variation of stomatal type in the families under studies. The main observations from the Table are highlighted below; from this data, the following four groups can be distinguished.
Woody plants
Aceraceae: In the Aceraceae, stomata are present only on the abaxial surfaces and are found at relatively high densities. The highest densities are observed in A. monspessulanum at 510 per mm2 and the lowest in A. hyrcanum at 366 per mm2. Only anomocytic stomata are found in this family; guard cell lengths range between 22.06 and 23.14 μm and the stomatal apertures are unusually narrow.
Anacardiaceae: In the Anacardiaceae stomata are absent from adaxial surfaces, with the exception of Pistacia atlantica which has stomata on both surfaces.
| Table 1: | Stomatal densities, Guard cell length for the adaxial and abaxial epidermis and type of stomata in dicotyledons (*=species showing a correlation between increased guard cell size and higher stomatal density: see discussion) |
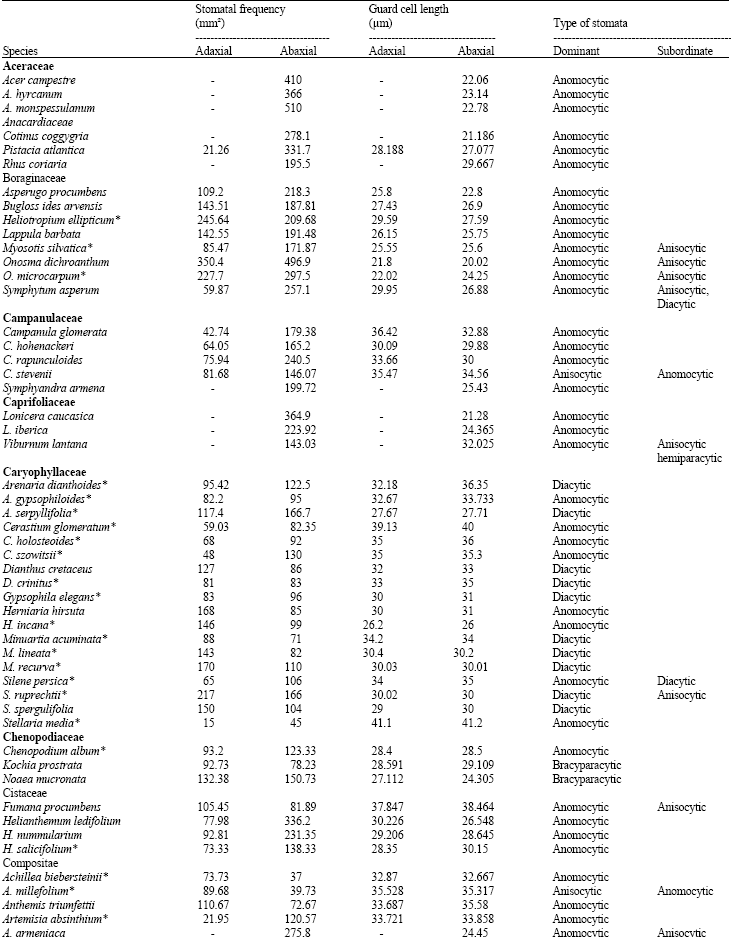 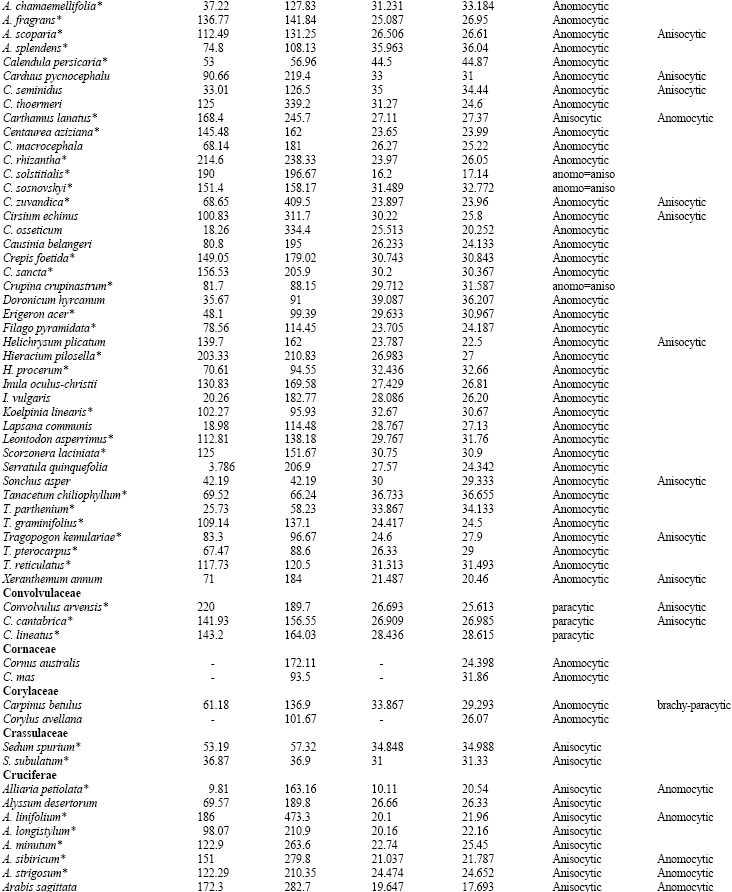 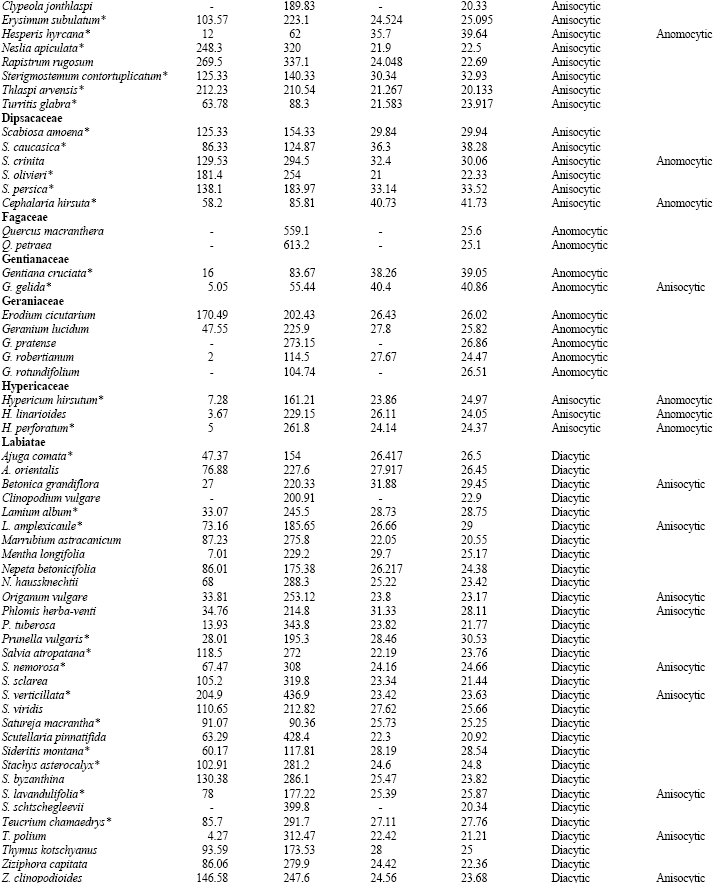 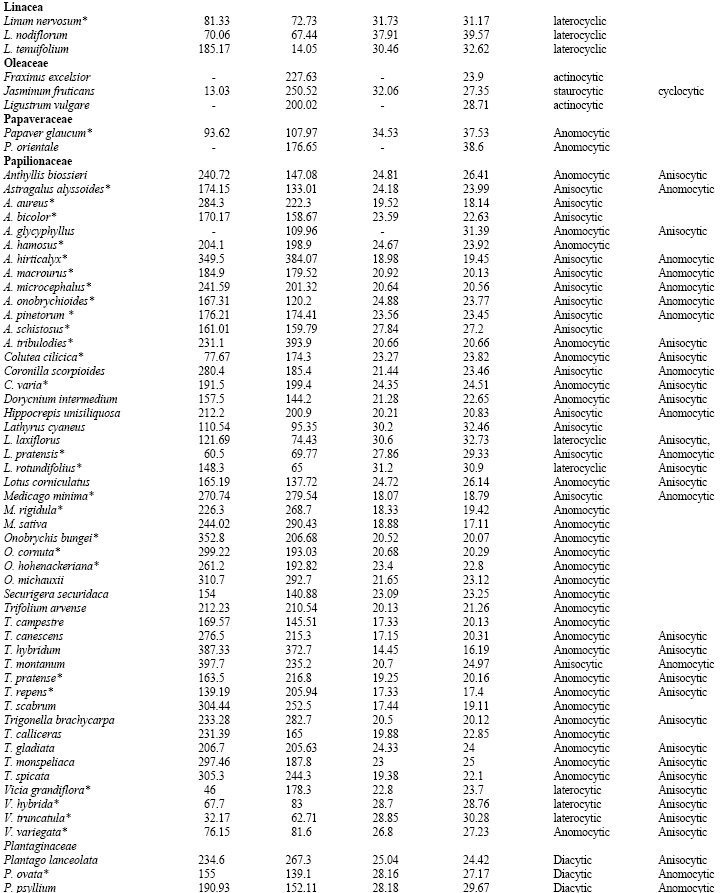   | |
Stomatal densities within the family range from 195 per mm2 for Rhus coriaria to 331 per mm2 for P. atlantica. The stomatal type is usually anomocytic and guard cells vary between 21.18 and 29.66 μm in length.
Caprifoliaceae: In the three species of Caprifoliaceae represented here, stomata are observed only on abaxial surfaces. The highest density is observed in Lonicera caucasica with an average of 364 per mm2. The dominant stomatal type is anomocytic. In Viburnum lantana, anisocytic and hemiparacytic stomata are also present as subordinate types. The guard cell length is between 21.28 and 32.02 μm.
Cornaceae: In the Cornaceae, stomata are found only on abaxial surfaces. The stomatal density is 93-172 per mm2. The dominant stomatal type is anomocytic. Guard cell length is between 24.39 and 31.86 μm.
Corylaceae: In the Corylaceae, only abaxial stomata are present, except in Carpinus betulus which has stomata on both surfaces; in this species, the average densities are 61 per mm2 and 136 per mm2 for adaxial and abaxial surfaces, respectively. Stomatal density is low in comparison with other woody plants. The dominant stomatal type is anomocytic, though in C. betulus brachyparacytic stomata are also present. Guard cell average length is 33.86 μm on adaxial surfaces and between 26.7 and 29.29 μm on abaxial surfaces for the two species studied.
Fagaceae: In the Fagaceae, stomata are observed only on abaxial surfaces. The stomatal density is high, averaging at 613 per mm2. The stomatal type is anomocytic and guard cell average length approximates to 25 μm.
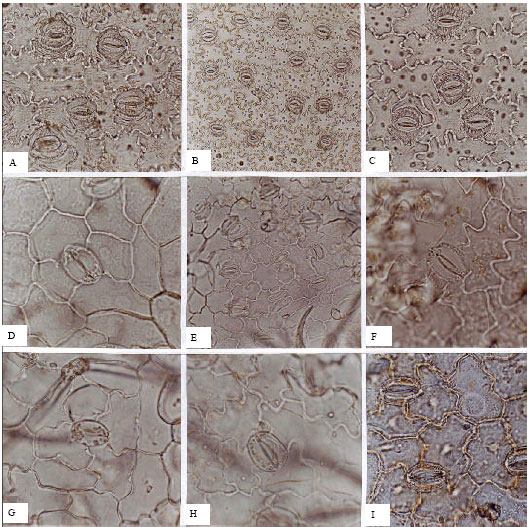
Oleaceae (Fig. 2A-I): In the Oleaceae, stomata are absent from adaxial surfaces, with the exception of Jasminum fruticans. Abaxial average densities are between 200-250 per mm2. Stomata are anomocytic, with actinocytic, staurocytic or cyclocytic arrangements in different species. Guard cell average length is 32.06 μm on adaxial surfaces and 23.9 μm on abaxial surfaces.
Rhamnaceae: In the Rhamnaceae, stomata are present only on abaxial surfaces, with the exception of Rhamnus pallasii. The highest stomatal density is observed in Paliurus spina-chiristi at 437 per mm2. Stomatal type is anomocytic. Guard cell lengths vary between 29.52 and 21.52 μm.
Rosaceae: In the Rosaceae, stomata are observed on both laminar surfaces in the genera Alchemilla, Geum, Potentilla and Spirea, whilst in Amygdalus, Cerasus, Cotoneaster, Prunus and Rosa, they are found only on abaxial surfaces.
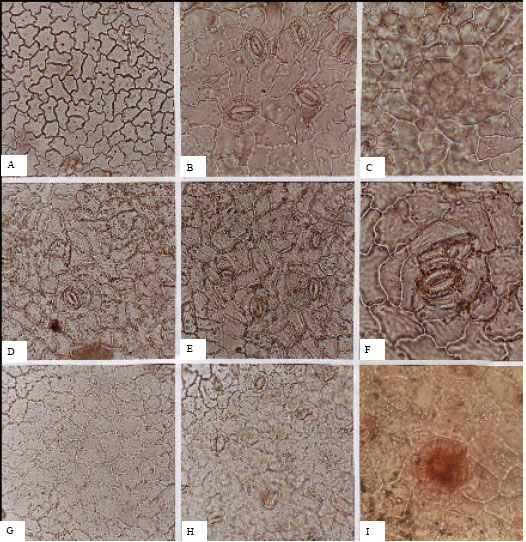 | |
| Fig. 2: | A-I, epidermis in superficial view of the Oleaceae; A-C, Fraxinus excelsior; A, adaxial; B, C, abaxial; C, glandular trichome; D-F, Jasminum fruticans; D, adaxial; E, F, abaxial; G-I, Ligustrum vulgare; G, adaxial; H, I, abaxial; I, glandular trichome; A, D, E, G, H, (x150); B, C, F, I, (x300) |
The abaxial stomatal densities are always higher than those of the adaxial surfaces; the highest density is observed in Prunus at an average of 408 per mm2. Stomatal type is anomocytic. Where stomata are present on both surfaces, the adaxial guard cells are larger (average 31.43 μm) than those of the abaxial surface (average 21.44 μm), though in the genus Spiraea, this is vice-versa.
Ulmaceae (Fig. 3A-G): In the Ulmaceae, stomata are present only on the abaxial surfaces. The stomatal densities are high, ranging between 280 per mm2 for Ulmus glabra and 742 per mm2 for Celtis caucasica. Stomata are anomocytic in C. caucasica whilst in the genus Ulmus, cells are cyclocytic. Guard cell lengths vary between 20.66 and 33.67 μm.
Herbaceous species with stomatal characteristics comparable to woody plants
Campanulaceae: In the Campanulaceae, with the exception of Symphyandra armena, stomata are present on both surfaces. Abaxial stomatal frequencies are usually higher than those of the adaxial surfaces; these differences can be up to four-fold, as in Campanula glomerata. The average stomatal densities are 42 per mm2 for adaxial surfaces and 240 per mm2 for abaxial surfaces. The dominant type of stomata is anomocytic, with the exception of C. stevenii where anisocytic patterning is dominant over the anomocytic arrangement. Guard cells on adaxial surfaces (average 36.42 μm) are longer than those of the abaxial surface (average 25.43 μm).
Cistaceae (Fig. 4A-I): In the Cistaceae, stomata are observed on both surfaces. The abaxial stomatal densities are greater than on those of the adaxial surfaces with the exception of Fumana procumbens.
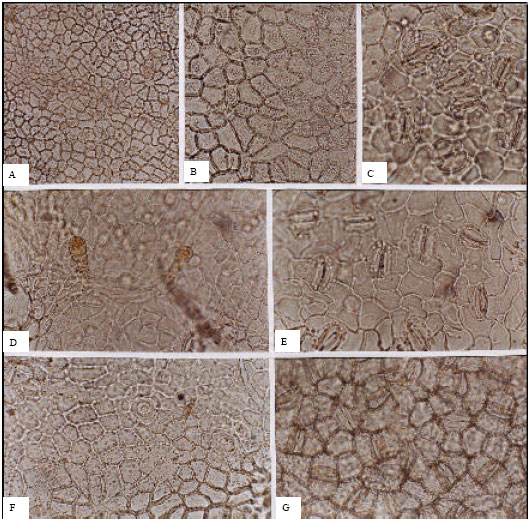 | |
| Fig. 3: | A-G, epidermis in surface view of the Ulmaceae; A-C, U. minor; A, B, adaxial; C, abaxial; D, E, U. glabra; D, adaxial; E, abaxial; F, G, Celtis caucasica; F, adaxial; G, abaxial; A, (x75); B, D, F, (x150); C, E, G, (x300) |
 | |
| Fig. 4: | A-I, epidermis in superficial veiw of the Cistaceae; A-C, Fumana procumbens; A, adaxial; B, C, abaxial; D-F, Helianthemum ledifolium; D, adaxial; E, F, abaxial; G, H, H. salicifolium; G, adaxial; H, abaxial; I, adaxial surface of H. nummularium; A, C, E, (x150); B, (x75); D, F-I, (x300) |
The density differences between adaxial and abaxial surfaces can be up to four-fold, as in Helianthemum ledifolium, which at an average of 336 per mm2 shows the highest density of the four species studied. The dominant stomatal type is anomocytic, though in F. procumbens anisocytic patterning is also observed. The guard cell lengths range between 26.54 and 38.46 μm for abaxial surfaces.
Geraniaceae: In the Geraniaceae, with the exceptions of G. pratense and G. rotundifolium, stomata are found on both surfaces. The abaxial stomatal densities are higher than those of adaxial surfaces. The difference between surfaces is considerable; higher densities on the abaxial side range between 104 and 273 per mm2. The stomatal type is anomocytic. Guard cells on adaxial surfaces (average 27.8 μm) are longer than those of the abaxial surfaces (average 24.47 μm).
Polygonaceae (Fig. 5A-I): In the Polygonaceae, stomata are present on both laminar surfaces. Abaxial stomatal densities (average 150 per mm2) are higher than those of the adaxial surfaces (average 59 per mm2). The dominant stomatal type is anisocytic, with anomocytic stomata present as a subordinate pattern. Guard cell lengths on adaxial surfaces (average 42.62 μm) are greater than abaxial guard cell average lengths (29.53 μm).
Primulaceae: In the Primulaceae, stomata are observed on both laminar surfaces, with higher abaxial stomatal densities except in Androsace villosa. Average densities for the four species studied are 18 per mm2 for adaxial surfaces and 117 per mm2 for abaxial surfaces.
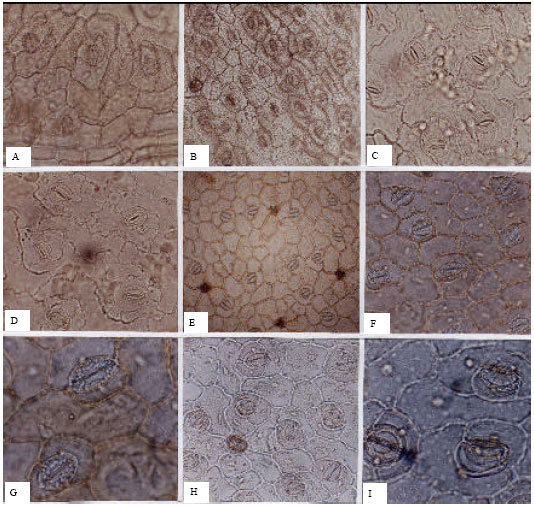 | |
| Fig. 5: | A-I, epidermis in superficial view of the Polygonaceae; A, B, Atraphaxis spinosa; A, adaxial; B, abaxial; C, D, Polygonum alpestre; C, adaxial; D, abaxial; E-G, Rumex acetosa; E, adaxial; F, G, abaxial; H, I, R. tuberosus; H, adaxial; I, abaxial; A, C, D, F, H, (x150); B, E, (x75); G, I, (x300) |
The differences in density between these surfaces can be up to six times, as in P. macrocalyx. Stomata are anomocytic throughout the family. Guard cell average length for the adaxial surfaces (42.58 μm) is greater than the abaxial average (27.85 μm).
Valerianaceae: In the Valerianaceae, stomata are present on both surfaces, with higher-abaxial stomatal densities (102 per mm2) than those of the adaxial surfaces (54 per mm2). The dominant stomatal type is anomocytic; anisocytic cells are also present as a subordinate category. Adaxial guard cell lengths are greater (average 37 μm) than abaxial cell lengths (average 26.83 μm).
Violaceae: In the Violaceae, stomata are present on both surfaces; with higher average abaxial (135 per mm2) than adaxial densities (15 per mm2). Stomatal type is anisocytic. The largest guard cells are always found on adaxial surfaces; cell lengths vary between 37.67 μm for adaxial and 27.06 μm for abaxial guard cells.
Families of herbaceous species with characters distinct from woody plants
Caryophyllaceae: In the Caryophyllaceae, special arrangements of stomata are evident in a genus- specific manner; e.g. in Herniaria, Minuartia and Silene (with the exception of S. persica) stomatal frequencies on adaxial surfaces are greater than those of abaxial surfaces, whilst in Arenaria and Cerastium these are vice-versa. In Dianthus, Minuartia, Arenaria (with the exception of A. gypsophiloides) and Silene (with the exception of S. persica) the dominant stomatal type is diacytic, whilst in Cerastium and Herniaria, anomocytic cells dominate. Usually abaxial stomata (average length 41.2 μm) are equal to or larger than the adaxial stomata (26.2 μm), e.g., in Cerastium and Silene, differences in guard cell lengths between adaxial and abaxial surfaces are minor. The size of stomata is similar on both surfaces in Stellaria media and in all representatives of the genus Minuartia (Zarinkamar, 2001).
Compositae: In the Compositae, stomata are present on both surfaces, with the exception of Artemisia armeniaca. The stomatal densities of on abaxial surfaces are usually higher than those of the adaxial surfaces, with some exceptions (Achillea and Anthemis). The species A. armeniaca, Carduus thoermeri, Centaurea zavandica, Cirsium osseticum, Crepis sancta, Hieracium pilosella, Inula vulgaris, Tanacetum graminifolius and Tragopogon reticulatus have the highest stomatal densities in their respective genera. C. zavandica, with an average of 409 per mm2 on abaxial surfaces, has the highest stomatal density of any member of the Compositae family included in the present study. In this family, only anomocytic and anisocytic stomatal forms are evident. In the genera Artemisia, Centaurea, Hieracium, Tragopogon and Tanacetum, the abaxial guard cells are equal to or larger than adaxial cells, whilst in Carduus, Cirsium and Inula this is vice-versa. Stomatal size is similar on both surfaces in Achillea millefolium and in the genus Crepis.
Convolvulaceae: Throughout this family, stomata are observed on the both surfaces of the leaf. Abaxial stomatal densities are higher than those of the adaxial surfaces, with the exception of Convolvulus arvensis. Density differences between both surfaces are minor. The dominant stomatal form is paracytic, whilst anisocytic cells are also observed in two of the three species studied. Guard cell sizes are similar on both surfaces (with the exception on C. arvensis). The guard cell lengths range between 25.61 and 28.61 μm; longer guard cells are associated with higher stomatal densities.
Crassulaceae: In the Crassulaceae, stomata are present on both leaf surfaces. A similar adaxial and abaxial stomatal size and density is usually observed; the two species studied have an average stomatal density of between 36 and 57 per mm2. Stomatal type is anisocytic and guard cell lengths are between 31.00 and 34.98 μm.
Cruciferae: In the Cruciferae, stomata are present on both leaf surfaces, with the exception of Clypeola jonthlaspi. The stomatal densities of abaxial leaf surfaces are usually greater than those of the adaxial surfaces; differences can be up to two-fold as in the genus Alyssum, or can be roughly equivalent, as in Thlaspi arvense. The highest density is observed in A. linifolium at 473 per mm2 on the abaxial surface and the lowest in Hesperis hyrcana, with 12 per mm2 on adaxial surfaces and 62 per mm2 on abaxial surfaces. The dominant stomatal type is anisocytic, with anomocytic cells also present in some species. Abaxial guard cell lengths (39.64 μm) are equal to or larger than adaxial guard cells (10.11 μm), with the exceptions of Arabis sagittata, Rapistrum rugosum and Thlaspi arvense.
Dipsacaceae (Fig. 6A-I): In the Dipsacaceae, stomata are observed on both surfaces, with greater abaxial than adaxial densities; differences between surfaces vary within the family. The highest, in Scabiosa crinita has an abaxial average of 294 per mm2, which is twice that of the adaxial surface (129.53 per mm2). The lowest difference between surfaces is seen in Cephalaria hirsuta (58 per mm2 adaxial and 85 per mm2 abaxial). The dominant stomatal type is anisocytic, whilst anomocytic cells are occasionally subordinate. Abaxial guard cells lengths are greater (average 41.73 μm) than adaxial guard cells (average 21 μm), with the exception of S. crinita. Longer guard cells are usually associated with higher stomatal frequencies.
Gentianaceae: In the Gentianaceae, stomata are present on both surfaces. The overall stomatal density is low, especially on adaxial surfaces. Average densities are between 5 and 16 per mm2 on adaxial surfaces and from 55 to 83 per mm2 on abaxial surfaces. The dominant stomatal type is anomocytic, whilst anisocytic cells are present as a subordinate category in G. gelida. Abaxial guard cells are longer (average 40.86 μm) than adaxial cells (average 38.26 μm).
Hypericaceae: In the family Hypericaceae, stomata are present on both leaf surfaces; abaxial are higher than adaxial stomatal densities, with most stomata present on abaxial surfaces (abaxial averages range between 161 and 261 per mm2). The dominant type is anisocytic, with anomocytic stomata also evident. Guard cell lengths on abaxial surfaces are greater than those of adaxial surfaces, with the exception of H. linarioides. Guard cell average lengths for the three species studied are between 23.86 and 26.11 μm (both of these are measurements of cells from adaxial surfaces).
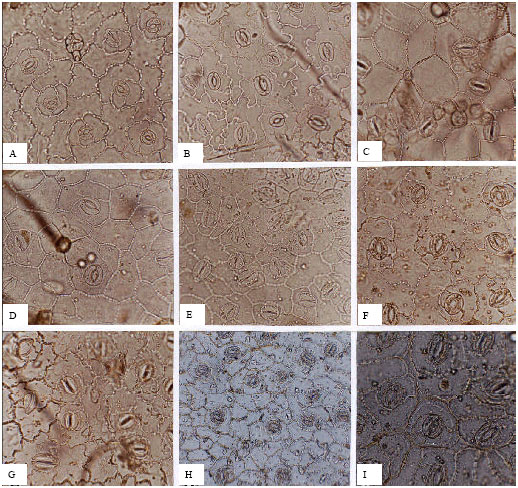 | |
| Fig. 6: | A-I, epidermis in superficial view of Scabiosa sp.; A, B, S. olivieri; A, adaxial; B, abaxial; C, adaxial epidermis of S. amoena; D, E, S. crinita; D, adaxial; E, abaxial; F, G, S. persica; F, adaxial; G, abaxial; H, I, S. caucasica; H, adaxial; I, abaxial; A-G, I, (x150); H, (x75) |
Papilionaceae (Fig. 7, A-H): The Papilionaceae, are well represented in this study, with 48 species examined. Stomata are observed on both surfaces throughout the family, with the exception of Astragalus glycyphyllus. Stomata show unusually wide variability and lack any genus-specific occurrence of a distinctive stomatal form; also the numbers of stomata per unit area are often higher on the adaxial surfaces. The highest density is observed in Trifolium (139 to 397 per mm2 on adaxial surfaces); and the lowest in the genus Vicia (32 per mm2 and 178 per mm2 on adaxial and abaxial surfaces, respectively). The dominant stomatal type in Astragalus is anisocytic; whilst in Trifolium and Trigonella anomocytic cells are most frequent. In Lathyrus and Vicia, laterocyclic cells are observed together with other two types. Guard cells are smaller in comparison with other families studied, ranging in length between 14.45 μm on the adaxial surface of Trifolium hybridum, to 32.73 μm on the abaxial surface of A. glycyphyllus.
Ranunculaceae: In the Ranunculaceae, stomata are found on both surfaces. The difference in density between adaxial and abaxial surfaces is minor, with the exceptions of Delphinium tuberosum and Thalictrum minus where stomata are completely absent from the adaxial surface. Generally abaxial stomatal densities are higher than those of the adaxial surfaces, except in Adonis aestivalis and Ranunculus strigillosus where this is vice-versa. The highest density is observed in T. minus (with 248 per mm2 on the abaxial surfaces) and the lowest in A. aestivalis (with 46 per mm2 on adaxial surfaces and 27 per mm2 on abaxial surfaces). Stomata are anomocytic in all species studied and abaxial guard cells are longer (average 61.33 μm) than adaxial cells (average 33.9 μm), except in the species A. aestivalis.
Families of herbaceous species whose stomatal characters differ greatly between genera of the same family and also within species of the same genus
Boraginaceae: In the Boraginaceae, stomata are observed on both leaf surfaces, though are always most abundant on the abaxial epidermis, with the exception of Heliotropium ellipticum.
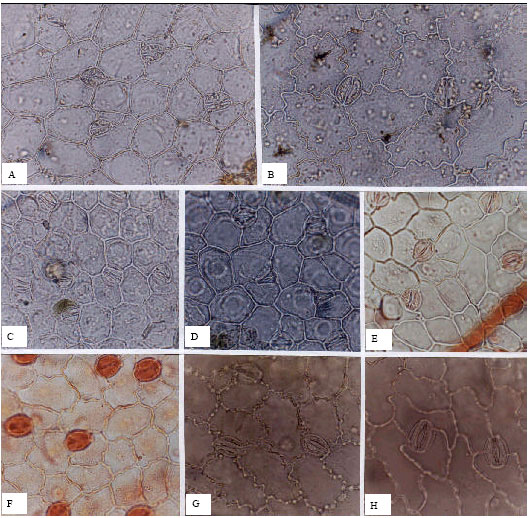 | |
| Fig. 7: | A-H, epidermis in superficial view of Trifolium sp.; A, C, E, adaxial; B, D, F, G, H, abaxial; A, B, T. pratense; C, D, T. hybridum; E, F, T. canescens; G, T. arvense; H, T. repens; A-H, (x300) |
In some species this difference is between two-to four-folds, as in Asperugo procumbens, Myosotis silvatica and Symphytum asperum. The highest stomatal densities are observed in Onosma dichroanthum (350 per mm2 on adaxial and 496 per mm2 on abaxial surfaces) and the lowest in M. silvatica (85 per mm2 on adaxial and 171 per mm2 on abaxial surfaces). The dominant stomatal type is anomocytic, with anisocytic cells present as a subordinate type in some species such as M. silvatica, O. dichroanthum and O. microcarpum. In S. asperum, diacytic cells are also observed together with the other two types. Guard cell average length on adaxial surfaces (29.95 μm) is greater than abaxial surfaces (20.2 μm), with the exception of O. microcarpum.
Chenopodiaceae: In the Chenopodiaceae, stomata are present on both surfaces, with greater abaxial than adaxial densities, except in Kochia prostrata. The differences between adaxial and abaxial surfaces are usually minor. Of the three species studied, densities are highest in Noaea mucronata (abaxial average of 150 per mm2) and lowest in K. prostrata (abaxial average of 78 per mm2). The dominant type of stomata in Chenopodium album is anomocytic, whilst in K. prostrata and N. mucronata bracyparacytic cells are most frequent. Guard cell average length is between 24.30 and 29.1 μm in these three species (Zarinkamar, 2006a).
Labiatae (Fig. 8A-J): In the Labiatae, stomata are found on both surfaces, except in Clinopodium vulgar and Stachys schtschegleevii, where adaxial stomata are completely absent. Abaxial stomatal densities are higher than those of the adaxial surfaces; the highest of these is observed in Salvia verticillata (436 per mm2) with significant differences between surfaces for all species studied. The dominant stomatal type is diacytic, with anisocytic forms present as a subordinate pattern.
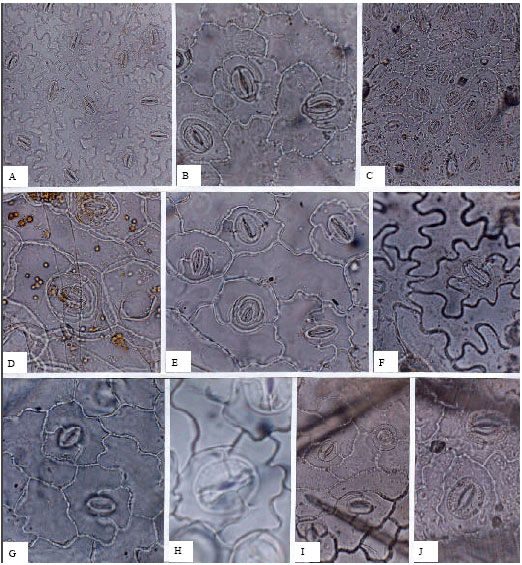 | |
| Fig. 8: | A-J, epidermis in superficial view of the Labiatae; A, abaxial epidermis of Lamium album; B, adaxial epidermis of Nepeta betonicifolia; C, abaxial epidermis of Ziziphora clinopodioides; D, E, Salvia atropatana; D, adaxial; E, abaxial; F, abaxial epidermis of Mentha longifolia; G, H, Scutellaria pinnatifida; G, adaxial; H, abaxial; I, J, Betonica grandiflora; I, adaxial; J, abaxial; A, C, I, (x150); B, D-G, J, (x300), H, (x750) |
Guard cell lengths on adaxial surfaces (average 31.88 μm) are usually equal to or larger than abaxial cell lengths (average 20.34 μm).
Linaceae: In the Linaceae, stomata are present on both surfaces. Adaxial stomatal densities (between 70 and 185 per mm2) are higher than those of abaxial surfaces. Density differences between surfaces are minor in Linum nervosum and L. nodiflorum, but vary greatly in L tenuifolium. Stomatal type is laterocyclic. Guard cells are large, averaging between 30.46 and 39.57 μm in length.
Papaveraceae: Two species of Papaveraceae occur in this study: of these, stomata are present on both surfaces in P. glaucum, whilst in P. orientale they are found only on the abaxial surfaces. The average densities for these two species are 93.62 per mm2 for adaxial surfaces and 176.65 per mm2 for abaxial surfaces. Stomatal type is anomocytic. Guard cell average length is 34.53 for adaxial cells and 38.6 μm for abaxial cells.
Plantaginaceae: In the Plantaginaceae, stomata are present on both surfaces in all species studied. Adaxial stomatal densities are higher than abaxial densities, with the exception of Plantago lanceolata. Densities range between 139 and 267 stomata per mm2 and differences between both surfaces are minor. The dominant stomatal type is diacytic; anomocytic and anisocytic forms are also present. Adaxial guard cells are often larger than abaxial cells; lengths range between 24.42 and 29.67 μm.
Rubiaceae: In the herbaceous members of the Rubiaceae studied here, stomata are present on both surfaces and abaxial densities are higher than those of the adaxial surfaces. Cruciata taurica and Galium odoratum have a shrubby growth form in this environment and in these species stomata are completely absent from the adaxial surface. The highest density of stomata is found on the abaxial leaf surfaces of G. verum with an average of 233 per mm2 and the difference between both surfaces is high. Stomata throughout this family are laterocyclic and are consistently large; the guard cell lengths range from 29.09 μm for abaxial cells, to 46.67 μm for adaxial cells.
Scrophulariaceae (Fig. 9A-I): In the Scrophulariaceae, stomata are found on both surfaces except in the genus Pedicularis. Abaxial stomatal densities are higher than those of the adaxial surface, except in Linaria kurdica. The highest overall average density is observed on the abaxial surfaces of species in the genus Pedicularis, at 353 per mm2. Density differences between both surfaces
show variability throughout this family, without any genus specific patterns. In most of the species studied, stomata are anomocytic; anisocytic cells are variably present as a subordinate type. Guard cell average lengths range from (abaxial) 21.86 μm to (adaxial) 39.60 μm.
Umbelliferae (Fig. 10A-H): In the Umbelliferae, most species, studied have stomata on both surfaces; the exceptions are Laser trilobum, Peucedanum caucasicum, Sanicula europaea and Seseli peucedanoides.
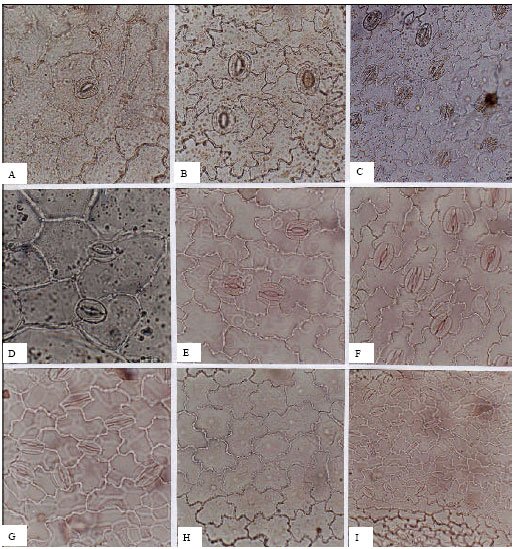 | |
| Fig. 9: | A-I, epidermis in superficial view of the Scrophulariaceae; A, B, Digitalis nervosa; A, adaxial; B, abaxial; C, abaxial epidermis of Veronica anagallis-aquatica; D, adaxial epidermis of V. polita; E, F, V. gentianoides; E, adaxial; F, abaxial; G, abaxial epidermis of Pedicularis sibthorpii; H, I, P. wilhelmsiana; H, adaxial; I, abaxial; A-C, E, F, H, (x150); D, G, (x300); I, (x75) |
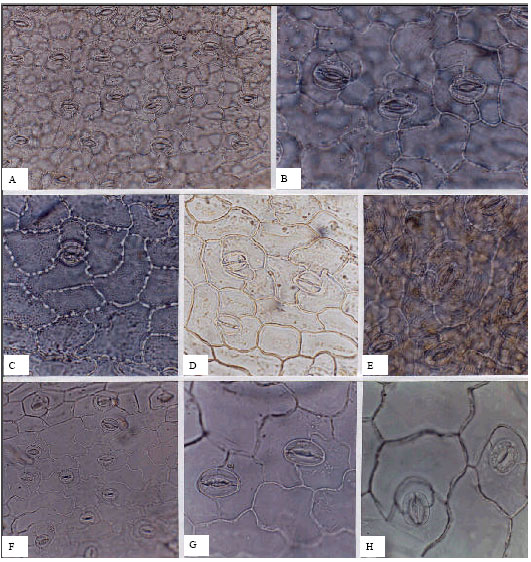 | |
| Fig. 10: | A-H, epidermis in superficial view of the Umbelliferae; A-E, G, abaxial; F, H, adaxial; A, B, Carum carvi; C, Bupleurum falcatum; D, B. geradii; E, Pimpinella tragium; F, G, Daucus broteri; H, Caucalis platycarpos; A, F, (x150); B-E, G, H, (x300) |
Abaxial stomatal densities are greater than those of the adaxial surfaces except in the genus Bupleurum. The highest observed abaxial density is found in Pimpinella aurea, at an average of 533 per mm2. Stomatal density differences between leaf surfaces illustrate the variability between species e.g., in Prangos ferulacea adaxial and abaxial surfaces are similar, whilst in Chaerophyllum aureum adaxial stomata are very sparse (4.44 per mm2) and the abaxial density is unusually high for the family (326.7 per mm2). Species in this family were found in a variety of different habitats and the types and sizes of stomata are variable both inter- and intra-specifically. For example in Chaerophyllum aureum, the commonest stomatal form recorded is the anisocytic pattern at (40%), followed by anomocytic (30%), then diacytic (20%) and last finally paracytic cells (10%); because of variation due to environmental influences, taxonomic classification is particularly difficult. Guard cell lengths range between 17.66 μm and 45.3 μm; both of these values represent abaxial stomata. In the Umbelliferae, stomatal types provide a valuable indicator for easier identification and classification at the species level.
As indicated, in Table, the types of stomata are highly variable between herbaceous species and so could be a valuable tool in informing their taxonomic classification.
Stomatal types in woody plants are mostly anomocytic; these plants show little variation compared with herbaceous species. Herbaceous plants include a much greater diversity of stomatal forms. These are, in decreasing order of occurrence, anomocytic, anisocytic, diacytic and paracytic. The family Umbelliferae encompass the greatest variety in stomatal form, whilst the least variation in form is seen in the families Ranunculaceae and Rubiaceae.
DISCUSSION
The numbers of stomata present on leaf surfaces vary widely amongst different species of plants. Generally the abaxial epidermis has higher numbers of stomata than the adaxial surface. According to Eames (1947), stomata on the leaves of woody plants are more frequent on the lower (abaxial) surfaces of leaves and the numbers per unit area tend to be greater than in herbaceous plants. Table 1 show that a higher density of stomata is observed in species where stomata are present only on one leaf surface.
Although the data generally confirm Eames' observations, some herbaceous species show a stomatal distribution similar to that of woody plants, due to an absence of stomata on their adaxial surfaces. This is clearly evident in, for example in Artemisia armeniaca (Compositae), Clypeola jonthlaspi (Cruciferae), Galium sp., Cruciata taurica (Rubiaceae) and Laser trilobum, Peucedanum caucasicum, Sanicula europaea and Seseli peucedanoides (Umbelliferae).
Interesting observations from this study include;
| • | In the Linacea, Plantaginaceae and the majority of Papilionaceae families, in contrary to the general features of herbaceous species studied, the adaxial stomatal densities are higher than those of the abaxial surfaces and |
| • | The similarity of stomatal sizes and densities on both surfaces of the Crassulaceae family. |
The highest recorded adaxial stomatal densities are found in Trifolium montanum (Papilionaceae), at 397 stomata per mm2. The highest densities for an abaxial surface are observed in Celtis caucasica (Ulmaceae) at 742 per mm2. The largest observed guard cell length is 65.5 μm in Adonis aestivalis (Ranunculaceae) and the smallest is 10.11 μm in Alliaria petiolata (Cruciferae); both of these measurements are from adaxial surfaces.
According to Willmer (1996), stomatal frequency and guard cell size are interdependent and the stomatal size usually decreases with increasing stomatal density. However the data presented in this paper shows that guard cells on lower leaf surfaces are equal to or smaller in size than those of the upper surfaces in some families. Table 1 show that Willmer’s observations apply generally to the woody plants, although some exceptions occur such as Alchemilla fluminea, Spiraea crenata and S. hypericifolia (Rosaceae). In these exceptions, in spite of low stomatal densities on the adaxial surfaces, the abaxial guard cells are larger than those of the adaxial surface.
In this study, an assessment of the validity of Willmer’s theory of the interdependence of guard cell size and stomatal density, demonstrates the following three phenomena:
| • | Willmer’s theory correlates generally to the stomatal patterning seen in first group (the woody plants) and in the following herbaceous families from group two of the results section; Campanulaceae, Cistaceae, Geraniaceae, Polygonaceae, Primulaceae, Valerianaceae and Violaceae. |
In many of these herbaceous species, there are major differences in stomatal density between adaxial and abaxial surfaces; these patterns are close to those observed in woody plants.
Willmer’s proposal is confirmed by the general observations from the first and second groups of families. The stomatal characters common to the second group species are as follows;
| • | Stomata are observed on both sides of leaves. |
| • | Abaxial stomatal densities are higher than adaxial densities. |
| • | The differences in density between both surfaces are high, tending towards greater stomatal densities on abaxial surfaces; this can be up to six-fold as in Primula macrocalyx (Primulaceae) or four-fold in Helianthemum ledifolium (Cistaceae); Campanula glomerata (Campanulaceae).and Geranium lucidum (Geraniaceae). |
| • | Stomatal type is anomocytic or anisocytic. |
| • | The largest guard cells are always found on the adaxial surfaces. |
| • | Stomatal size usually decreases with increasing stomatal frequency. |
| • | The third group of plant families demonstrates a patterning phenomenon that is in contrast to Willmer’s findings, characterised by a correlation between increased guard cell size and increased stomatal density. This group includes the following families; Caryophyllaceae, Compositae, (with the exception of the genera Inula, Carduus and Cirsium), Convolvulaceae, Cruciferae, Dipsacaceae, Gentianaceae, Hypericaceae, Papilionaceae and Ranunculaceae. In most of these species, the stomatal density differences between adaxial and abaxial surfaces are minor: |
Common stomata characters in this group, in contrast with Willmer’s hypothesis, are as follows;
| • | Stomata are present on both surfaces. |
| • | Abaxial stomatal densities are higher than on adaxial densities, except some genera from the Caryophyllaceae and the majority of Papilionaceae. |
| • | The density differences between both surfaces are minor, except in the Hypericaceae and Gentianaceae. |
| • | The abaxial guard cells are larger than adaxial cells. |
| • | Longer guard cells are associated with higher stomatal densities. |
| • | The types of stomata are variable within families. |
| • | The fourth group of plant families demonstrate variable phenomena within the same genus; some species show characteristics of group II, in line with Willmer’s theory, whilst other species show the opposing pattern as in group III above. This category includes genera from the following families: Boraginaceae, Chenopodiaceae, Labiatae, Linaceae, Papaveraceae, Plantaginaceae, Rubiaceae, Scrophulariaceae and Umbelliferae. |
Previous studies of various herbaceous species have shown that if stomata are found on both surfaces of the leaf, then usually the adaxial guard cells are larger than those of the abaxial surface. The data in Table 1 demonstrate that, contrary to Willmer’s observations, in certain herbaceous species a higher stomatal density is associated with increased guard cell size. These are indicated in the Table 1; with an asterisk. In almost all of these, the density differences between adaxial and abaxial surfaces are minor. This suggests that the relationship between density and guard cell size is strongly related to density differences and distribution of stomata over both leaf surfaces in herbaceous plants.
The adaptive significance of the presence of stomata over one or both leaf surfaces is unclear. According to Parkhurst (1978), herbaceous plants increase CO2 uptake by reducing the length of the CO2 diffusion pathway to the mesophyll. It may be that in herbaceous species, the efficiency of photosynthesis is potentially higher than that of woody plants, because of the existence of stomata on both surfaces of the leaf.
Although the rate of photosynthesis in woody plants may be less per unit area than in herbaceous species, the absence of stomata on the adaxial surfaces and the variable densities of these abaxial stomata, suggest a greater capacity for response and adaptation to environmental conditions, as seen in the xerophytes growth form.
REFERENCES
- Dilcher, D.L., 1974. Approaches to the identification of angiosperm leaf remains. Bot. Rev., 40: 1-157.
Direct Link - Retallk, G.J., 2001. 300 million year record of atmospheric carbon dioxide from fossil plant cuticles. Nature, 411: 287-290.
CrossRefDirect Link - Zarinkamar, F., 2006. Density, size and distribution of stomata in different monocotyledons. Pak. J. Biol. Sci., 9: 1650-1659.
CrossRefDirect Link