
Tony Hadibarata
Department of Applied Bioscience, Faculty of Agriculture, Ehime University 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566, Japan
Sanro Tachibana
Department of Applied Bioscience, Faculty of Agriculture, Ehime University 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566, Japan
Kazutaka Itoh
Department of Applied Bioscience, Faculty of Agriculture, Ehime University 3-5-7 Tarumi, Matsuyama, Ehime, 790-8566, Japan
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 11 | Page No.: 1804-1810







ABSTRACT
The degradation of n-eicosane by Trichoderma sp. S019, a fungus collected from soil with extensive degradative ability on an agar medium containing n-eicosane, was demonstrated in liquid medium and in soil. Maximal degradation (77%) was obtained when Trichoderma sp. S019 was incubated for 30 days after the addition of 0.1 mM of n-eicosane to the liquid medium while the highest rate of degradation (40%) was obtained in soil with the addition of 1.5% Trichoderma sp. S019. Furthermore, the degradation of n-eicosane was affected by the addition of a carbon source, the addition of a nitrogen source and agitation. n-Eicosane was indeed degraded by Trichoderma sp. S019 because nonadecanoic acid, n-octadecane, hexadecanoic acid, oleic acid and stearic acid, considered to be the intermediates in the biodegradation of n-eicosane, were detected among the reaction products.
PDF Abstract XML References Citation
How to cite this article
Tony Hadibarata, Sanro Tachibana and Kazutaka Itoh, 2007. Biodegradation of n-Eicosane by Fungi Screened from Nature. Pakistan Journal of Biological Sciences, 10: 1804-1810.
DOI: 10.3923/pjbs.2007.1804.1810
URL: https://scialert.net/abstract/?doi=pjbs.2007.1804.1810
DOI: 10.3923/pjbs.2007.1804.1810
URL: https://scialert.net/abstract/?doi=pjbs.2007.1804.1810
INTRODUCTION
Alkanes are an acyclic saturated hydrocarbon commonly found in petroleum-contaminated environments and the biodegradation of these compounds has been well studied So et al. (2003). Crude oil is primarily composed of hundreds of different hydrocarbon molecules, mainly alkanes from C1 to C40 straight chains, C6 to C8 branched-chains, cyclohexanes, aromatics and compounds containing sulphur, nitrogen and oxygen (Stafford et al., 1982; Prince, 1993). The majority of molecules in crude oil spills and refined products are biodegradable and they will eventually disappear from the environment as they are utilized by microbes (Prince, 1993). These are generally considered or be weakly biodegradable compounds compared with most of the organic molecules involved in the biological carbon cycle (Bertrand et al., 1989). Several studies have shown that aromatic compounds have a greater resistance to microbial degradation than n-alkanes which are considered to be the most readily degradable hydrocarbons (Ehrhardt and Blumer, 1972; Higgins and Gilbert, 1978).
The ability of microorganisms to degrade hydrocarbons and facilitate their mineralization by forming more labile organic compounds through the breakdown of intramolecular bonds has been extensively studied. Microbial degradation is a rate-limiting factor in many biogeochemical cycles. As a result, microorganisms have contributed to the development of different bioremediation technologies (Aeckersberg et al., 1991; Arora and Gill, 2001; Madsen, 1991).
Aeckersberg et al. (1991) reported that a degradation mechanism of alkane in which the chain lengths of alkane substrates had a pronounced effect on the total cellular fatty acid composition. An alkane with an odd number of C atoms yielded predominantly fatty acids with an even number of C atoms and vice versa. Alkane was altered by an odd number carbon during its transformation to fatty acid. Phelps et al. (2001) reported using stable isotope-labeled hydrocarbons as substrates to help delineate the pathway of anaerobic degradation and examine the initial mechanisms of attack of compounds such as benzene, naphthalene and alkane.
In the present report, we describe the results of an investigation of the degradation of n-eicosane by Trichodermasp. S019, a fungus collected from soil and capable of degrading phenanthrene. The catabolic pathway for n-eicosane was also examined.
MATERIALS AND METHODS
Chemicals: n-Eicosane, nonadecanic acid, n-octadecane, hexadecanoic acid and stearic acid were purchased from Tokyo Chemical Industry Co. Ltd. (Tokyo, Japan). Malt extract and polypeptone were acquired from Difco (Detroit, USA). Thin Layer Chromatography (TLC) aluminium sheets (Silica gel 60 F254, 20x20 cm) were obtained from Merk (Darmstadt, Germany). The silica gel used for column chromatography (Wakogel S-1) and all other chemicals were purchased from Wako Pure Chemical Industry Co. Ltd. (Osaka, Japan).
Fungi and medium: The one hydrocarbonoclastic fungus Trichoderma sp. S019 was originally isolated from soil. The fungus is able to grow on alkanes, particularly on n-eicosane, as its sole source of carbon and energy. Trichoderma sp. S019 was isolated from soil at the Campus of the Faculty of Agriculture, Ehime University. A 5 mm disk of selected fungi was aseptically inoculated into a 100 mL Erlenmeyer flask containing 20 mL of Mineral Salt Broth (MSB) containing (in g L-1 distilled water): glucose (10), KH2PO4 (2), MgSO4.7H2O (0.5), CaCl2.2H2O (0.1), Ammonium tartrate (0.2) and trace elements (10 mL) as described by Arora and Gill (2001). Each flask was incubated for 15 or 30 days at 25°C in the dark. After autoclaving for 20 min at 120°C, 1 mM of n-eisosane dissolved in 1 mL of dimethylformamyde containing 100 μL of tween 80 was added to the flask. Control experiments were performed by incubating MSB medium with the test alkane without the inoculum. All assays were conducted in triplicate. The carbon concentration-dependence of n-eicosane’s degradation was investigated on media containing glucose in the concentration range 4-10% of liquid medium. The nitrogen concentration-dependence of n-eicosane’s degradation was investigated on media containing polypeptone in the concentration range 4-10% of liquid medium. Agitation at 120 rpm was used to enhance the degradation of n-eicosane. All media were sterilized by autoclaving at 120°C for 20 min.
Soil treatment: Soil samples were collected from the A Horizon (0-20 cm from soil surface) with pH 5 in Matsuyama City. The soil samples were air dried and sieved through 3 mm mesh. Soil was autoclaved at 120°C for 20 min to eliminate the effect of microorganisms in it. A solution of n-eicosane dissolved in water containing dimethylformamyde and tween 80 was added to the soil and mixed well. To ensure the homogeneity of the treatments, the soil was then sieved again through 3 mm mesh. The final concentration of n-eicosane in the soil was measured at 1 ppm. Trichoderma sp. S019 was grown in the mineral salt broth medium for 2 weeks. The fungal suspension was applied in droplets to the soil surface and then mixed thoroughly with a sterile spatula on a clean bench.
Analytical methods: After incubation of the liquid media, the culture broth was blended with ethyl acetate and acidified with 1N HCl. The filtrate (liquid medium) and residue (fungal body) were separated by filtration and then the liquid medium and fungal body were extracted with ethyl acetate, respectively. Each extract was combined and purified by column chromatography using 200 mL of dichlorometane:methanol (19:1). With this method, all substrates initially present in the liquid medium were recovered. The extracts were concentrated and analyzed by gas chromatography-mass spectrometry (GC-MS Shimadzu QP-5050). The amount of substrate was determined using 4-chlorobiphenyl as an internal standard. GC-MS was used with the following conditions: column 30 m in length and 0.25 mm in diameter, helium pressure 100 kPa. The temperature program was started at 80°C, held for 2 min, raised from 80°C to 200° at 20°C/min, then to 260°C at 7.5°C/min, then held for 4 minutes. The flow rate was 1.5 mL min-1, interface temperature was 260°C and injection volume was 1 μL.
After 15 and 30 days of incubation, the soil (30 g dry weight equivalent) was placed in a Soxhlet apparatus and refluxed for 16 hours with 300 mL of dichloromethane. After cooling, the solvent was evaporated to obtain a small amount of extract. Column chromatography was performed in order to purify the extract using 200 mL of dichloromethane:methanol (19:1). GC was then conducted as described earlier.
Detection of metabolites: The culture medium was prepared as described above. After inoculation of the medium with Trichoderma sp. S019, the culture was pre-incubated by standing for 7 days at 25°C in the dark. n-Eicosane (5.65 mg) dissolved in 100 μL of N, N-Dimethylformamide (DMF) and 10 μL of Tween 80 (1% solution) was added to each culture medium as described above. The incubation was conducted for 7-30 days at 25°C in the dark. The extracts were purified using silica gel column chromatography by successive elution with n-hexane, n-hexane and ethyl acetate (1:1 v/v), ethyl acetate and ethanol. As authentic compounds, nonadecanoic acid, hexadecanoic acid and stearic acid could not be detected directly by GC-MS, so an analytical derivatization procedure was used. These compounds were subjected to TMS methods. TLC and GC analyses were initially performed to indicate the presence of different intermediates in the pathway for the degradation of n-eicosane. GC-MS was then carried out to conclusively prove the presence of these intermediates.
Following the vacuum drying of each eluate (100 μL) in a vial, N, O-bis-trimethylsilyl acetamide (40 μL), pyridine (40 μL) and trimethylcholorilane (20 μL) were added, respectively. Trimethylsilylation of the eluate was conducted for 10 min at 80°C without contact with moisture. The trimethylsilyl (TMS) derivatives of each eluate were analyzed by gas chromatography (GC) using Shimadzu GC-17 equipped with a TC-1 capillary column (30 mx0.25 mm) ID 0.25 μm using a gradient of 60°C for 2 min, raised to 150°C at 15°C/min, then raised to 300°C at 25°C/min and maintained at 300°C for 6 min. Injector and interface temperatures were 260°C. In order to confirm the intermediates/metabolites of n-eicosane’s degradation and to determine the degradation pathway, a Gas chromatograph-mass spectrophotometer, Shimadzu QP5050, was used in this experiment. The conditions for the GC-MS analysis consisted of operating a detector at 1.3eV, scan intervals of 1s and a mass range of 50-500.
RESULTS
Investigation of the degradation of n-eicosane in a liquid medium: Trichoderma sp. S019 degraded 77 and 46% of n-eicosane at 0.1 mM and 1 mM in 30 days, respectively. There was a simultaneous decrease in the concentration of substrate during the course of the incubation. Trichoderma sp. S019 gave the highest total degradation at a concentration of 0.1 mM (Table 1).
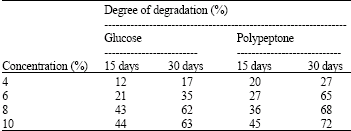
Investigation of the effect of carbon and nitrogen sources on the degradation of n-eicosane in a liquid medium: n-Eicosane was readily degraded in the fungal cultures in the present study. Trichoderma sp. S019 degraded 17, 35, 62 and 63% of n-eicosane at 4, 6, 8 and 10% glucose in 30 days, respectively. The maximum degradation was 63% when Trichoderma sp. S019 was incubated in the culture with 10% glucose in 30 days. Addition of polypeptone was performed in order to know the effect of a nitrogen source on the degradation of the compound. Trichoderma sp. S019 degraded 57, 65, 68 and 72% of n-eicosane at 4, 6, 8 and 10% polypeptone in 30 days. The highest rate of degradation was 72% when Trichoderma sp. S019 was incubated in the culture with 10% polypeptone for 30 days (Table 2).
The effect of agitation on the degradation of n-eicosane in shake-flask cultures in the dark was examined. Trichoderma sp. S019 degraded 55 and 61% of n-eicosane at 80 and 120 rpm, respectively. The maximal degradation rate was 81% when Trichodermasp. S019 was incubated in the culture with shaking at 120 rpm for 30 days (Table 3).
Investigation of the degradation of n-eicosane in soil: The rate of degradation of n-eicosane in soil was above 15% for 30 days of incubation with Trichodermasp. S019. Trichoderma sp. S019 degraded 19, 28 and 40% of n-eicosane in soil on treatment with 0.5, 1.0 and 1.5% of fungi at 1 ppm in 30 days, respectively. There was a simultaneous decrease in the concentration of substrate during the incubation period. Trichoderma sp. S019 gave the highest degradation rate in soil on treatment with 1.5% fungi (Table 4).
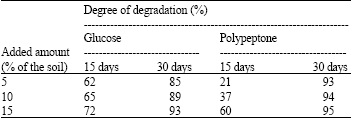
Addition of carbon and nitrogen sources had been conducted in order to increase the rate of degradation of n-eicosane in soil by Trichoderma sp. S019 degraded 85, 89 and 93% of n-eicosane at 5, 10 and 15% glucose in 30 days, respectively. The maximum degradation was 93% when Trichoderma sp. S019 was incubated in a soil with 15% glucose for 30 days. Addition of polypeptone was performed in order to know the effect of a nitrogen source on the degradation of the substrate. Trichoderma sp. S019 degraded 93, 94 and 95% of n-eicosane at 5, 10 and 15% polypeptone in 30 days. The highest rate of degradation was 95% when Trichoderma sp. S019 was incubated in soil with 15% polypeptone for 30 days (Table 5).
| Table 1: | Degradation of n-eicosane by Trichoderma sp. S019 in liquid medium |
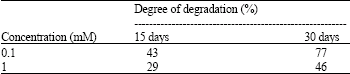 | |
| Table 2: | Effect of nutrients on degradation of n-eicosane by Trichoderma sp. S019 in liquid medium |
 | |
| Table 3: | Effect of agitation on degradation of n-eicosane by Trichoderma sp. S019 in liquid medium |
 | |
| Table 4: | Degradation of n-eicosane by Trichoderma sp. S019 in soil |
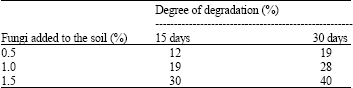 | |
| Table 5: | Effect of nutrients on degradation of n-eicosane by Trichoderma sp. S019 in soil |
 | |
Identification of metabolites: To confirm the degradation of n-eicosane in the liquid medium with fungi, the identification of intermediates was conducted. According to the degradation pathway for alkane with bacterium shown by So et al. (2003), alkane was subterminally carboxylated to form an intermediate and two adjacent carbon atoms were then eliminated to form a fatty acid one carbon shorter than the original alkane. Identification of these compounds in the extracts using the method described in the experimental section was conducted. In the Total Ion Chromatograph (TIC) profile, the retention times of these compounds coincided with those of the authentic compounds. The Mass Spectrum (MS) of the trimethylsilyl (TMS) ether of these compounds also coincided with that of the authentic TMS ether of the compounds.
The cellular fatty acid composition of Trichoderma sp. S019 was clearly affected by the chain length of the alkane substrate. Trichoderma sp. S019, grown in MSB liquid medium with n-eicosane for 15 days, was able to mineralize n-eicosane to form nonadecanoic acid (C-odd) and n-octadecane. Two peaks having a GC retention time at 8.9 and 9.3 min were detected. The GC retention time at 8.9 min, GC-MS spectrum of the TMS-derivative M+ at m/z 312 and fragment ions at m/z 297 (M+-15), corresponding to the respective sequential losses of methyl (-CH3), were identical to those of authentic nonadecanoic acid. Furthermore, the appearance of a peak at m/z 254 indicated that an alkane with less C atoms then the parent compound was also produced (Fig. 1).
Several additional n-eicosane metabolites were also detected by GC-MS analysis of the ethyl acetate-extractable soluble metabolites. Two peaks having a GC retention time at 9.3 and 10.1 min were detected. The peak at 9.3 min refers to authentic hexadecanoic acid. MS analysis of the hexadecanoic acid produced from nonadecanoic acid gave an apparent molecular ion (M+) at m/z 328 for TMS derivatives and an apparent loss of (M+-15) at m/z 313, representing a probable loss of methyl (-CH3) as well as expected fragmentation at m/z 73 (Fig. 2). The other peak having a GC retention time at 10.1 minutes exhibited an apparent molecular ion (M+) at m/z 354 and fragment ion at [M+-15] at m/z 339, representing a probable loss of methyl (-CH3.). This mass spectral fragmentation pattern suggested that this metabolite was oleic acid (Fig. 2).
An anaerobic oxidation of n-eicosane had been also conducted by Trichodermasp. S019 in MSB medium. Trichoderma sp. S019 appeared to anaerobically oxidize alkane carbon to fatty acid, as evidenced by the influence of alkane carbon chain length on its cellular fatty acid composition.
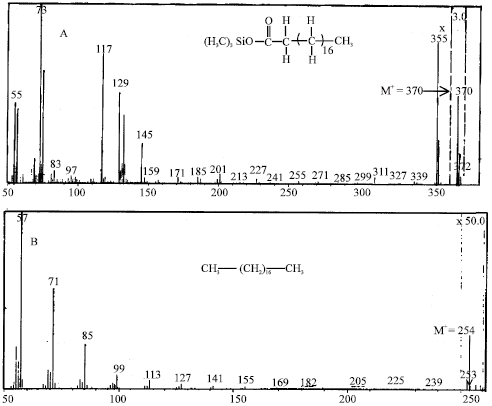 | |
| Fig. 1: | Mass spectra of the reaction product from the biodegradation of n-eicosane by Trichoderma sp. S019 screened from nature. (A) mass spectrum of the trimethylsilyl (TMS) ether of nonadecanoic acid produced in the biodegradation of n-eicosane, (B) mass spectrum of n-octadecane produced in the biodegradation of n-eicosane |
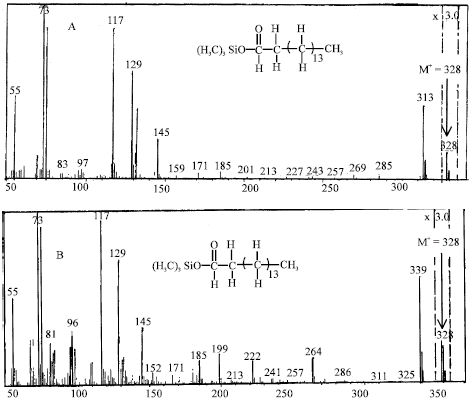 | |
| Fig. 2: | Mass spectra of the trimethylsilyl (TMS) ether of the reaction product from the biodegradation of nonadecanoic acid by Trichoderma sp. S019 screened from nature. (A) mass spectrum of hexadecanoic acid produced by the biodegradation of nonadecanoic acid, (B) mass spectrum of oleic acid produced by the biodegradation of nonadecanoic acid |
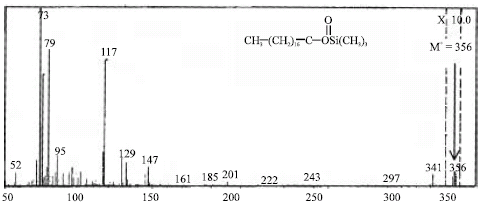 | |
| Fig. 3: | Mass spectrum of the reaction product from the biodegradation of n-eicosane in liquid culture without CO2 by Trichoderma sp. S019 screened from nature |
Several n-eicosane metabolites were also detected by GC-MS analyses of the ethyl acetate-extractable soluble metabolites. One major peak having a GC retention time at 10 min refers to authentic stearic acid. Mass spectral fragmentation of the stearic acid produced from n-eicosane gave an apparent molecular ion [M+] at m/z 356 for a TMS derivative and an apparent loss of [M+-15] at m/z 341 corresponding to the respective sequential loss of methyl (-CH3), as well as the expected fragment ion at m/z 73. Based on the data obtained above, the compound should be stearic acid (Fig. 3).
DISCUSSION
Trichoderma sp. S019 utilized n-eicosane as a sole carbon source for growth. In the present experiment, when n-eicosane was provided as the carbon source, an usually long period of 30 days incubation was required to attain a very low level of n-eicosane. One of the reasons for this could be that n-eicosane was used in very high concentrations coupled with it being the only exogenous carbon source provided. Al-Hadhrami et al. (1996) showed that addition of an alternative carbon and nutrient source, such as mollase, increased repiration and n-alkane degradation in synthetic seawater. We suggested that the addition of carbon and nitrogen sources such as glucose and polypeptone, increased the metabolic activity and thereby stimulated the production of enzymes effective in alkane’s degradation. This result is also similar to that of Khleifat (2006) who found that the rate of degradation of a substrate increased after addition of a carbon source to the medium. Carbon sources at 0.2%, except fructose, allowed phenol degradation to proceed faster at ranges between 1.4-fold and 3-fold higher than that of the control and in all cases the growth of the microorganism was in accordance with the standard microbial batch growth culture.
Similar results were obtained for the degradation of n-eicosane in soil, in that the addition of nutrients increased the rate of biodegradation. An increase in the concentration of nutrients increased the rate of degradation by Trichoderma sp. S019. The available nutrients were rapidly assimilated by soil microbes, thus depleting the reserve of nutrients. Consequently, the objective of adding nutrients to the soil samples was to restore the availability of essential nutrients. The degradation rate was also affected by the size of the fungal population added to the soil. Several researchers have recently described an increase in microbial activity and rate of biodegradation following the addition of nutrients (Radwan et al., 2000; Rahman et al., 2003).
Consistent with reports of the deleterious effect of agitation on enzyme production, the agitation of growing shake-flask cultures on a rotary shaker at 120 rpm resulted in the formation of mycelial pellets. This result is similar to that of Collina et al. (2005) who found the oxygen concentration is directly dependent on the air flow rate. On the one hand, stirring increases the contact between the reagents (substrate, oxygen and biomass), thus enhancing mass transfer and, as a consequence, the biodegradation. On the other hand, agitation increases the contact surface between the slurry phase and the gas phase in the bioreactor, thus enhancing the loss by volatilization of oxygen and substrates and decreasing the biodegradation rate.
The strains oxidize alkanes to fatty acids and incorporate them into cellular lipids, a metabolic process observed in many aerobic alkane–degrading organisms.
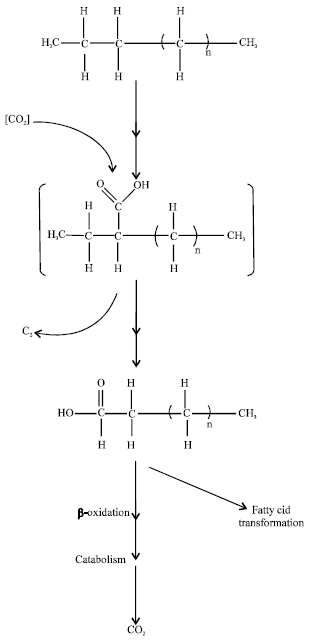 | |
| Fig. 4: | A Proposed pathway for the degradation of n-eicosane by Trichoderma sp. S019 with reference to the results of So et al. (2003) |
The carbon numbers of fatty acids also correlate with those of the alkane substrate. A C-odd alkane yielded predominantly C-even fatty acids and a C-even alkane yielded C-odd fatty acids. The initial attack on alkane involves a novel mechanism in which an odd-numbered carbon was added to or removed from the alkane chain (Aeckersberg et al., 1991; Doumenq et al., 2001). In this study, we directly confirmed that n-eicosane was oxidized to nonadecanoic acid by Trichodermasp. S019. It was suggested that C-even alkanes are indeed transformed to C-odd fatty acids by Trichoderma sp. S019.
We also conclude that n-eicosane is first transformed to a fatty acid which is one carbon shorter (nonadecanoic acid) by the addition of an exogenous carbon atom and by the removal of two terminal carbon atoms from the alkane chain. As summarized in Fig. 4, we propose that the initial attack of n-eicosane involves subterminal carboxylation at the atom carbon number 3 (C-3) position of the alkyl chain. The adjacent carbon atom located at C-1 was then removed to yield a fatty acid. This fatty acid contains a carboxyl group exogenously added by the carboxylation reaction, making it one carbon shorter than the parent alkane. This fatty acid then undergoes β-oxidation to form an n-saturated fatty acid which can be further β-oxidized and eventually mineralized to CO2 (Kunau et al., 1995; Mao et al., 1995). Alternatively, this fatty acid can undergo transformations such as chain elongation and methylation to form other fatty acids (Rahman et al., 2003).
A different mechanism behind the degradation of n-eicosane in the liquid medium without CO2 was also clarified. The reaction appears to occur subterminally at C-3 of the alkane carbon chain and its followed by the removal of two adjacent terminal carbon atoms to form a fatty acid. n-Eicosane reacted at the C-3 position and then the two adjacent carbon atoms at C-1 and C-2 were eliminated. A fatty acid (stearic acid) was subsequently formed which was two carbons shorter that the original alkane.
ACKNOWLEDGMENTS
We thank Mr. Djarwanto, Forest Product Technology Research and Development Center, Ministry of Forestry, Bogor, Indonesia for identification of the fungus Trichoderma sp. S019.
REFERENCES
- Aeckersberg, F., F.A. Rainey and F. Widdel, 1998. Growth, natural relationships, cellular fatty acids and metabolic adaptation of sulfate-reducing bacteria that utilize long-chain alkanes under anoxic conditions. Arch. Microbiol., 170: 361-369.
CrossRefDirect Link - Al-Hadhrami, M.N., H.M. Lappin-Scott and P.J. Fisher, 1996. Effects of addition of organic carbon source on bacterial respiration and n-alkane biodegradation of Omami crude oil. Mar. Poll. Bul., 32: 351-357.
Direct Link - Arora, D.S. and P.K. Gill, 2001. Comparison of two assay procedures for lignin peroxidase. Enzyme Microb. Tech., 28: 602-605.
CrossRef - Collina, E., G. Bestetti, P. Di Gennaro, A. Franzetti, F. Gugliersi, M. Lasagni and D. Pitea, 2005. Naphthalene biodegradation kinetics in an aerobic slurry-phase bioreactor. Environ. Int., 31: 167-171.
CrossRef - Doumenq, P., E. Aries, L. Asia, M. Acquaviva, J. Artaud, M. Gilewick, G. Mille and J. C. Bertrand, 2001. Influence of n-alkanes on petroleum on fatty acid composition of a hydrocarbonoclastic bacterium: Marinobacter hydrocarbonoclastic strain 617. Chemosphere, 44: 519-528.
CrossRef - Ehrhardt, M. and M. Blumer, 1972. The source identification of marine hydrocarbons by gas chromatograph. Environ. Poll., 3: 179-194.
CrossRefDirect Link - Khleifat, K.M., 2006. Biodegradation of phenol by Ewingella Americana: Effect of carbon starvation and some growth conditions. Process Biochem., 41: 2010-2016.
CrossRef - Kunau, W.H., V. Dommes and H. Schultz, 1995. β-Oxidation of fatty acids in mitocondria, peroxisomes and bacteria: A century of continued progress. Prog. Lipid Res., 34: 267-342.
CrossRef - Mao, L.F., C. Chu, M.J. Luo, A. Simon, A.S. Abbas and H. Schultz, 1995. Mitochondrial β-oxidation of 2-methyl fatty acids in rat liver. Arch. Biochem. Biophys., 321: 221-228.
CrossRefDirect Link - Phelps, C.D., X. Zhang and L.Y. Young, 2001. Use of stale isotop to identify benzoate as a metabolite of benzene degradation in a sulfidogenic consortium. Environ. Microbiol., 3: 600-603.
Direct Link - Prince, R.C., 1993. Petroleum spill bioremediation in marine environment. Crit. Rev. Microbiol., 19: 217-242.
PubMed - Radwan, S.S., D. Al-Mailem, I. El-Nemr and S. Salamah, 2000. Enhanced remediation of hydrocarbon contaminated desert soil fertilized with organic carbons. Int. Biodeter. Biodegr., 46: 129-132.
CrossRef - Rahman, K.S.M., T.J. Rahman, Y. Kourkoutas, I. Petsas, R. Marchant and I.M. Banat, 2003. Enhanced bioremediation of n-alkane in petroleum sludge using bacterial consortium amended with rhamnolipid and micronutrients. Bioresour. Technol., 90: 159-168.
CrossRefDirect Link