
G. Mosayebi
Department of Histology, School of Medicine, Arak University of Medical Sciences, Arak, Iran
A. Ghazavi
Department of Histology, School of Medicine, Arak University of Medical Sciences, Arak, Iran
H. Salehi
Department of Histology, School of Medicine, Arak University of Medical Sciences, Arak, Iran
M. A. Payani
Department of Histology, School of Medicine, Arak University of Medical Sciences, Arak, Iran
M. R. Khazaei
Department of Histology, School of Medicine, Arak University of Medical Sciences, Arak, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 11 | Page No.: 1790-1796







ABSTRACT
Sesame oil was evaluated in the treatment of in C57BL/6 mice. It has profound anti-inflammatory activity and been traditionally used to treat inflammatory disorders. EAE was induced by immunization of 6-8 week old mice with MOG35-55 with complete Freunds adjuvant. Therapy with sesame oil was started on day 3 before the immunization. Total Antioxidant Capacity (TAC) was assessed by Ferric Reducing-antioxidant Power (FRAP) method. Nitric Oxide (NO) production was also estimated by Griess reaction. For histological analysis, mice brain was harvested and sections were stained with Hematoxylin-Eosin. After daily intraperitoneal dosage the sesame oil significantly reduced the clinical symptoms in C57BL/6 mice with EAE. Also, treated mice displayed a significantly delayed disease onset compared with control mice. Sesame oil significantly increased TAC, but it’s effect on serum nitrite production was not significant. Typical brain leukocyte infiltration was observed in control mice compared with treated mice. Present results suggest for the first time that sesame oil therapy may be effective in the prevention of symptomatic EAE. This resistance to encephalomyelitis may be associated with inhibition of oxidative stress.
PDF Abstract XML References Citation
How to cite this article
G. Mosayebi, A. Ghazavi, H. Salehi, M. A. Payani and M. R. Khazaei, 2007. Effect of Sesame Oil on the Inhibition of Experimental Autoimmune Encephalomyelitis in C57BL/6 Mice. Pakistan Journal of Biological Sciences, 10: 1790-1796.
DOI: 10.3923/pjbs.2007.1790.1796
URL: https://scialert.net/abstract/?doi=pjbs.2007.1790.1796
DOI: 10.3923/pjbs.2007.1790.1796
URL: https://scialert.net/abstract/?doi=pjbs.2007.1790.1796
INTRODUCTION
Multiple Sclerosis (MS) is an inflammatory demyelinating disease of the Central Nervous System (CNS) that afflicts more than million people worldwide (Dean, 1994). There is no medical treatment available so far than can cure MS (Clegg and Bryant, 2001; Donoghue and Greenlees, 2000). Experimental Autoimmune Encephalomyelitis (EAE) is the most useful model of MS (Gold et al., 2000). Many clinical and pathological features of EAE show close similarity to the human disease MS; therefore EAE has been commonly used as a model system to study the mechanism of MS pathogenesis and to test the efficacy of potential therapeutic agents for the treatment of MS (Owens and Sriram, 1995). The pathogenesis of EAE comprises immunization with encephalitogenic myelin antigens in the presence of adjuvant, presentation of such antigens to T cells, migration of activated T cells to the CNS and development of inflammation and/or demyelination upon recognition of the same antigens (Owens and Sriram, 1995). An important aspect of the pathogenesis of EAE, with potential for therapeutic manipulation, is the role of free radicals such as Nitric Oxide (NO) in the inflammatory process (Staykova et al., 2006; Teixeira et al., 2005; Dalton and Wittmer, 2005; Staykova et al., 2005; Nazliel et al., 2002; Murphy et al., 2002). Thus a scavenger of Reactive Oxygen Species (ROS) is expected to prevent these free radical mediated EAE. There is increasing interest in the protective biological function of natural antioxidants contained in dietary plants, which are candidates for the prevention of oxidative damage (Aniya et al., 2005).
Sesame seeds (Sesamum indicum, Linn, Pedaliaceae) have long been categorized as a traditional health food in Iran and other Asian countries. It has also been traditionally used to treat inflammatory disorders. A number of lipid-soluble antioxidants have been isolated from sesame seeds, including sesaminol, sesamolin, P1 and pinoresinol; sesame oil has been found to contain considerable amounts (up to 1.5%) of the sesame lignans, sesamin and sesamolin (Kang et al., 1998).
The lignans present in sesame oil are thought to be responsible for its antioxidant and anti-inflammatory properties. However, no previous study has examined the use of sesame oil in the treatment of MS or other TH1 cell-mediated inflammatory diseases of the CNS. In the present study, we examined the effect of sesame oil on the day of onset and severity of EAE, NO production, total antioxidant capacity and leukocyte infiltration into the brain.
MATERIALS AND METHODS
Mice: Inbred male C57BL/6 mice were obtained from the Pasture Institute of Iran, at 6-8 weeks age. The mice (weighting about 20±2 g) were randomly divided into two groups, each with 10 mice. The mice were housed at the animal house at Immunologic research center in accordance with all Institutional Ethical and Health guidelines.
Induction of experimental autoimmune encephalomyelitis (EAE): Male C57BL/6 mice (6-8 weeks age and 20±2 g weight) were inoculated s.c. in the flank with 0.1 mL of an emulsion containing 200 μg of the encephalitogenic peptide MOG35-55 (MEVGWYRSPFSRVVHLYRNGK; Diapharm Ltd, Russia) and equal volume of complete Freund’s adjuvant (Sigma, St. Louis, MO) supplemented with 4 mg mL-1 Mycobacterium tuberculosis H37RA (Difco, Detroit, MI, USA). Mice were them injected intraperitoneally with 400 ng of pertussis toxin (Sigma) on the day of immunization and 2 days later (Costa et al., 2003).
Sesame oil treatment: All mice were fed for the duration of the experiment with tap water and mouse fodder. In the experimental group (n = 10), mice were given intraperitoneally sesame oil (4 mL/kg/day). Control mice (n = 10) were given intraperitoneally Phosphate-buffered Saline (PBS), (4 mL/kg/day). Sesame oil and PBS injection were started on day 3 before the immunization.
Clinical evaluation of EAE: Following the encephalitogenic challenge, mice were monitored daily and neurological impairment was scored on an arbitrary clinical score as follows: 0, no clinical sign; 1, partial loss of tail tonicity; 2, complete loss of tail tonicity; 3, flaccid tail and abnormal gait; 4, hind leg paralysis; 5, hind leg paralysis with hind body paresis; 6, hind and foreleg paralysis; 7, moribund or death. The relapse was defined when a mouse developed an increase of the clinical score (more than 1) accompanied by weight loss. Under recommendation of the animal ethics committee, mice were killed on 25 day post immunization (Mosayebi et al., 2006).
Brain histology: On day 25 (sacrifice day) the brain were taken from sesame oil-treated and control EAE mice, fixed in 10% buffered formalin and embedded in paraffin. Sections (5 μM thick) were cut and mononuclear cell infiltration were visualized by Hematoxylin-Eosin (H and E). Briefly, H and E staining sections were air-dried, fixed in 10% formaldehyde, dehydrated, stained for 5 min with hematoxylin, then 2 min with eosin (Sigma), then dehydrated and mounted in Permount. Semi-quantitative histological evaluation was performed using the following scored based on the severity of inflammation: 0, no inflammation; 1, cellular infiltrates only in the perivascular areas; 2, mild cellular infiltrates in parenchyma (1-10/section); 3, moderate cellular infiltrates in parenchyma (11-100/section); 4, marked cellular infiltrates in parenchyma (>100/section) (Okuda et al., 2002).
Total antioxidant capacity (TAC): TAC was measured by FRAP (Ferric reducing antioxidant power) method (Benzie and Strain, 1996). In brief, three reagents were used: 1) sodium acetate, acetic acid buffer (pH 3.6) ; 2) 10 mmol L-1 solution of 2, 4,6-tripyridyl-s-triazine in a 40 mmol L-1 solution of hydrochloric acid (Sigma, St. Louis, MO) and 3) 20 mmol L-1 solution of ferric chloride hexahydrate prepared in deionized water. The FRAP reagent was prepared daily with 25 mL of reagent one, 2.5 mL reagent two and three that were heated to 37°C before using. Sample was added to reagent in cuvettes with an autosampler and then read on a spectrophotometer at 593 nm at four minutes. FeSO4 was used as a standard and antioxidant power expressed as μmol L-1 FRAP.
Nitric oxide (NO) production: NO production was measured by determining the increase in nitrite concentration in serum using the Griess reaction adapted to microtitre plates (Kayhan et al., 2003). Briefly, 100 μL serum was mixed with 100 μL Griess reagent (equal volumes of 1% [weight/vol] sulfanilamide in 5% [vol/vol] phosphoric acid and 0.1% [weight/vol] naphtylethylenediamine-HCl), in flat bottomed 96 well microtiter plates and incubate at room temperature for 10 min and then the absorbance at 550 nm was measured using a microplate reader (Stat-Fax 2001). The amount of nitrite in the samples (in micro molar units) was calculated from a sodium nitrite standard curve freshly prepared in distilled water. Nitrite concentration expressed as μmol L-1.
Statistical analysis: Statistical analysis was performed with the Student’s t-test for the mean maximal score, mean onset day, histological score and level of nitrite and TAC in serum. p<0.05 was considered significant.
RESULTS
In the current study, we investigated the effect of Sesame Oil (SO) administration on EAE. Immunization with 150 μg of MOG35-55 peptide induced severe EAE in control mice. Figure 1 shows the effect of SO on body weight (A) and clinical scores (B) in SO-treated and control EAE mice. Significant improvement of weight was observed in the group treated with SO. Also, SO-treatedmice displayed a significantly delayed disease onset (day 14±1) compared with control mice (day 10±1).
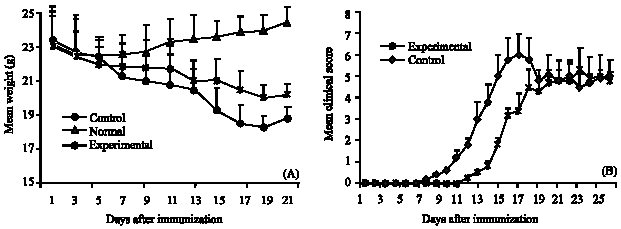 | |
| Fig. 1: | Mean body weight (A) and clinical course of MOG35-55-induced EAE (B) in C57BL/6 mice administrated sesame oil or vehicle. Mice were monitored daily for symptom and scored as described in materials and methods. Values are means and standard deviations for the 10 mice tested daily in each group |
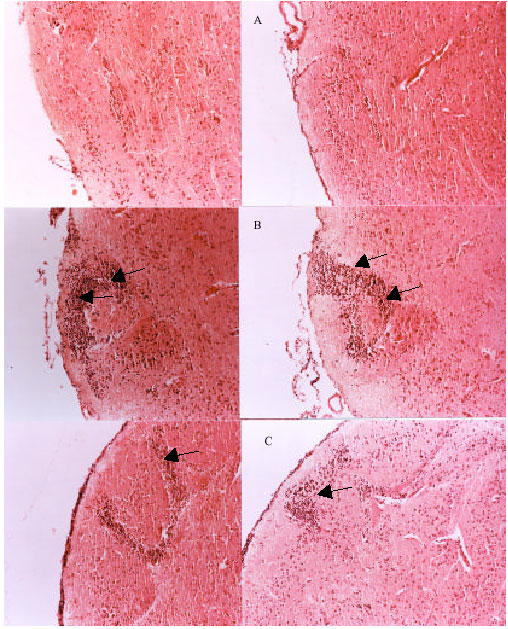 | |
| Fig. 2: | Brain sections from normal mice (A) and MOG-induced EAE mice treated with PBS (B) and sesame oil (C) were stained with Hematoxylin and Eosin (H and E) for leukocyte infiltration. Leukocyte infiltrates are less evident in the sesame oil-treated mice (C) in comparison with control mice (B) |
| Table 1: | Clinical features of EAE in control and sesame oil treated mice |
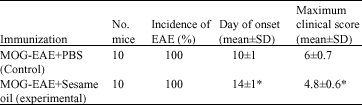 | |
| *Significantly different than control, p<0.05 | |
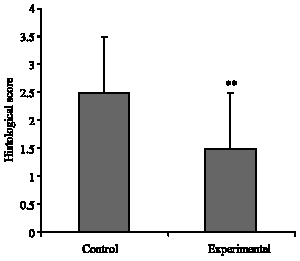 | |
| Fig. 3: | Histological score of the brain of C57BL/6 mice with MOG35-55-induced EAE at day 25 after immunization. Twelve sections cut at different levels of brain (randomly) of each mice (10 mice per group) were evaluated and scored as described in section materials and methods. *: significantly different than the control group, p<0.05 |
Also, the disease was less severe in the SO-treated group. All animals in SO-treated and control groups developed clinical EAE. There was significant reduction in maximum clinical score of EAE following SO administration (Table 1).
Histological findings: To examine the effect of sesame oil on amelioration of EAE, we histologically analyzed the brain of control and SO-treated EAE mice. For this study, ten mice in each group were administrated with either SO (4 mL/kg/day) or PBS from on day 3 before the first MOG immunization up to the sacrificed day (day 25 after immunization). As shown in Fig. 2, the brain of treated mice with SO showed less cell infiltrates (C) than that in control mice (B). To evaluate quantitatively the inflammatory lesions, the mean lesion area was calculated as described in materials and methods. Figure 3 show that a significant reduction in the mean lesion area in sesame oil-treated group is demonstrated compared to that in control group (p<0.05).
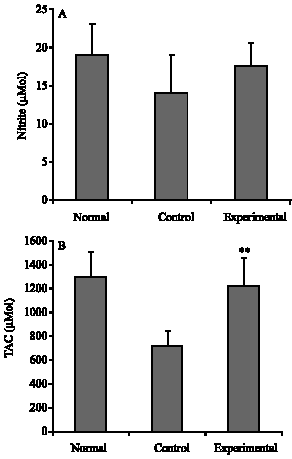 | |
| Fig. 4: | Concentration of nitrite (A) and Total antioxidant capacity (TAC) (B) in sera of normal C57BL/6 mice and both sesame oil-treated (Experimental) and control EAE mice. Data are mean±SD (n = 10). *p<0.05 compared with other groups |
TAC and NO concentration: Sera obtained from normal mice and both control and SO-treated EAE mice at day 25 after immunization were analyzed for Nitrite oxide production and TAC (Fig. 4A and B, respectively). The results show that the level of nitrite in SO-treated mice was more than that of control mice, although this difference was not significant. The mean TAC of the SO-treated group was significantly more than that of control group (p<0.01).
DISCUSSION
In this study, we have demonstrated a protective effect of Sesame Oil (SO) on EAE induced in C57BL/6 mice. SO significantly delayed the disease onset of EAE and reduced the maximum clinical scores. The mechanism by which SO exhibits its observed beneficial effect is still unknown, but some studies showed that SO has potent anti-inflammatory and antioxidant effects (Chavali et al., 2001; Akimoto et al., 1993). Hsu et al. (2004) showed that, SO protected rats against oxidative stress and hepatic injure after Cecal Ligation and Puncture (CLP) -induce chronic septic shock. In the other study, they observed that SO ameliorated hepatic and renal damage in a dose-dependent manner and increased animal survival and decreased lipid peroxide concentration in sera of lipopolysaccharide-treated rats (Hsu and Liu, 2002). Also, sesame oil consumption influences beneficially the blood glucose, lipid peroxidation and antioxidant levels in streptozotocin diabetic rats (Ramesh et al., 2005).
Earlier studies found evidence of lipid peroxidation in the CSF (Hunter et al., 1985) and in the plasma of patients with MS (Naidoo and Knapp, 1992). Some studies in patients with MS have shown increased free radical activity and deficiencies in antioxidant enzymes compared with healthy controls (LeVine, 1992). Free radicals such as Nitric Oxide (NO) may be important in the pathogenesis of EAE and MS. NO is an important mediator implicated in pathophysiological processes. Various investigations have indicated the high expression of iNOS and excessive amount of NO production in the pathogenesis of murine inflammatory CNS demyelination (MacMicking et al., 1992; Okuda et al., 1995). Furthermore, an excessive increase of NO production was found in both the CSF and the peripheral blood of MS patients (Yuceyar et al., 2001; Sherman et al., 1992). In contrast, protective role has been demonstrated for NO in EAE (Willenborg et al., 1999). NO has been shown to inhibit the cytotoxic effects of superoxide and hydrogen peroxide to neuronal cells. NO may also react with peroxyl radicals such as lipid peroxyl radicals and inhibit free radical chain reactions such as lipid peroxidation. Lipid peroxidation play an important role in pathogenesis of EAE and MS (Hogg, 1995).
Because of the pathogenic role of oxygen and nitrogen free radicals in MS pathology, antioxidants might prevent free radical-mediated tissue destruction and inhibit some of the early pro-inflammatory events, such as T cell trafficking into the CNS, that lead to inflammation and tissue destruction in EAE and MS.
Thus, with consider of increase stress oxidative in multiple sclerosis and EAE, SO as an antioxidant agent may suppress clinical score of EAE by affecting nitric oxide production and Total Antioxidant Capacity (TAC) of serum. Present results showed that SO significantly increases TAC in serum of EAE mice but serum nitrite production was not significantly different between the control and SO-treated in EAE mice. Hsu et al. (2005) reported that SO significantly inhibited the production of NO in rats with CLP-induced sepsis. In the other study, they showed that serum nitrite concentrations were not significantly different between the saline- and SO-treated in lipopolysaccharide challenge rats. Also, they investigated the effect of SO on oxidative stress-associated renal injury induced by lipopolysaccharide in rats. SO attenuated lipopolysaccharide-induced renal injury, decreased lipid peroxidation, increased the activities of superoxide dismutase, catalase, reduced hydroxyl radical generation and nitric oxide production (Hsu et al., 2005). Hou et al. (2003) showed that sesamin and sesamolin significantly inhibited NO production, iNOS mRNA and protein expression in LPS-stimulated BV-2 cells.
These conflict results can be related to the species of animal, route and dosage of SO administration.
Also, histological examination of brain tissues demonstrated reduced levels of infiltration of leukocytes in the SO-treated EAE mice. SO has anti-inflammatory effect and it has been suggested that SO or constituents of SO induces growth arrest and apoptosis of cancer. Recently, reported that SO, a vegetable oil enriched with n-6 polyunsaturated fatty acids attenuated the growth and metastasis of EL4 lymphoma (Salem, 2005).
In conclusion, this study shows, for the firs time, that SO reduces severity of disease and infiltration leukocytes to brain in EAE mice. However, the findings suggest that sesame oil might be potentially useful for the treatment of MS.
ACKNOWLEDGMENT
This research was supported by Research Council of Arak University of Medical Sciences, Project No. 182.
REFERENCES
- Akimoto, K., Y. Kitagawa, T. Akamatsu, N. Hirose, M. Sugano, S. Shimizu and H. Yamada, 1993. Protective effects of sesamin against liver damage caused by alcohol or carbon tetrachloride in rodents. Ann. Nutr. Metab., 37: 218-224.
Direct Link - Aniya, Y., T. Koyama, C. Miyagi, M. Miyahira, C. Inomata, S. Kinoshita and T. Ichiba, 2005. Free radical scavenging and hepatoprotective actions of the medicinal herb, Crassocephalum crepidioides from the Okinawa Islands. Biol. Pharm. Bull., 28: 19-23.
CrossRefDirect Link - Benzie, I.F.F. and J.J. Strain, 1996. The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": The FRAP assay. Anal. Biochem., 239: 70-76.
CrossRefPubMedDirect Link - Chavali, S.R., T. Utsunomiya and R.A. Forse, 2001. Increased survival after cecal ligation and puncture in mice consuming diets enriched with sesame seed oil. Crit. Care Med., 29: 140-143.
Direct Link - Clegg, A. and J. Bryant, 2001. Immunomodulatory drugs for multiple sclerosis: A systematic review of clinical and cost effectiveness. Exp. Opin. Pharmacother., 2: 623-639.
CrossRefDirect Link - Dalton, D.K. and S. Wittmer, 2005. Nitric-oxide-dependent and independent mechanisms of protection from CNS inflammation during Th1-mediated autoimmunity: Evidence from EAE in iNOS KO mice. J. Neuroimmunol., 160: 110-121.
CrossRef - Gold, R., H.P. Hartung and K.V. Toyka, 2000. Animal models for autoimmune demyelinating disorders of the nervous system. Mol. Med. Today, 6: 88-91.
CrossRef - Hou, R.C.W., H.L. Chen, J.T.C. Tzen and K.C.G. Jeng, 2003. Effect of sesame antioxidants on LPS-induced NO production by BV2 microglial cells. Neuroreport, 14: 1815-1819.
Direct Link - Hsu, D.Z. and M.Y. Liu, 2002. Sesame oil attenuates multiple organ failure and increases survival rate during endotoxemia in rats. Crit. Care Med., 30: 1859-1862.
Direct Link - Hsu, D.Z., Y.H. Li, S.P. Chien and M.Y. Liu, 2004. . Effects of sesame oil on oxidative stress and hepatic injury after cecal ligation and puncture in rats. Shock, 21: 466-469.
Direct Link - Hsu, D.Z., S.B. Su, S.P. Chien, P.J. Chiang, Y.H. Li, Y.J. Lo and M.Y. Liu, 2005. Effect of sesame oil on oxidative-stress-associated renal injury in endotoxemic rats: Involvement of nitric oxide and proinflammatory cytokines. Shock, 24: 276-280.
Direct Link - Kayhan, B., R. Aharoni and R. Arnon, 2003. Glatiramer acetate (Copaxone) regulates nitric oxide and related cytokine secretion in experimental autoimmune encephalomyelitis. Immunol. Lett., 88: 185-192.
CrossRef - MacMicking, J.D., D.O. Willenborg, M.J. Weidemann, K.A. Rockett and W.B. Cowden, 1992. Elevated secretion of reactive nitrogen and oxygen intermediates by inflammatory leukocytes in hyperacute experimental autoimmune encephalomyelitis: Enhancement by the soluble products of encephalitogenic T cells. J. Exp. Med., 176: 303-307.
Direct Link - Murphy, P., A. Sharp, J. Shin, V. Gavrilyuk and C. Dello Russo et al., 2002. Suppressive effects of ansamycins on inducible nitric oxide synthase expression and the development of experimental autoimmune encephalomyelitis. J. Neurosci. Res., 67: 461-470.
Direct Link - Nazliel, B., D. Taskiran, C. Irkec, F.Z. Kutay and S. Pogun, 2002. Serum nitric oxide metabolites in patients with multiple sclerosis. J. Clin. Neurosci., 9: 530-532.
Direct Link - Okuda, Y., Y. Nakatsuji, H. Fujimura, H. Esumi, T. Ogura, T. Yanagihara and S. Sakoda, 1995. Expression of the inducible isoform of nitric oxide synthase in the central nervous system of mice correlates with the severity of actively induced experimental allergic encephalomyelitis. J. Neuroimmunol., 62: 103-112.
Direct Link - Okuda, Y., M. Okuda and C.C. Bernard, 2002. Gender does not influence the susceptibility of C57BL/6 mice to develop chronic experimental autoimmune encephalomyelitis induced by myelin oligodendrocyte glycoprotein. Immunol. Lett., 81: 25-29.
CrossRef - Ramesh, B., R. Saravanan and K.V. Pugalendi, 2005. Influence of sesame oil on blood glucose, lipid peroxidation and antioxidant status in streptozotocin diabetic rats. J. Med. Food, 8: 377-381.
Direct Link - Salem, M.L., 2005. Systemic treatment with n-6 polyunsaturated fatty acids attenuates EL4 thymoma growth and metastasis through enhancing specific and non-specific anti-tumor cytolytic activities and production of TH1 cytokines. Int. Immunopharmacol., 5: 947-960.
Direct Link - Sherman, M.P., J.M. Griscavage and L.J. Ignarro, 1992. Nitric oxide-mediated neuronal injury in multiple sclerosis. Med. Hypotheses, 39: 143-146.
Direct Link - Staykova, M.A., S.A. Fordham, G.J. Bartell, W.B. Cowden and D.O. Willenborg, 2006. Nitric oxide contributes to the resistance of young SJL/J mice to experimental autoimmune encephalomyelitis. J. Neuroimmunol., 176: 1-8.
CrossRef - Teixeira, S.A., A.A. Varriano, A.A. Dias, R. Martins Porto and M.N. Muscara, 2005. Nitric oxide synthase activity and endogenous inhibitors in rats recovered from allergic encephalomyelitis. Mem. Inst. Oswaldo Cruz., 1: 25-27.
CrossRef - Yuceyar, N., D. Taskiran and A. Sagduyu, 2001. Serum and cerebrospinal fluid nitrite and nitrate levels in relapsing-remitting and secondary progressive multiple sclerosis patients. Clin. Neurol. Neurosurg., 103: 206-211.
CrossRef