
S. Qanbari
Research Institute of Physiology and Biotechnology, University of Zanjan, P.O. Box, 313, Zanjan, Iran
M.P. Eskandari Nasab
Department of Animal Science, University of Zanjan, P.O. Box, 313, Zanjan, Iran
R. Osfoori
Agricultural Biotechnology Research Institute of Iran, Karaj, Iran
A. Hagh Nazari
Research Institute of Physiology and Biotechnology, University of Zanjan, P.O. Box, 313, Zanjan, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 10 | Page No.: 1632-1638







ABSTRACT
We report the genetic analysis of 100 individuals of an elite breeding flock of Afshari sheep with a selected set of eighteen microsatellite markers. A full characterization of this set of eighteen loci was carried out generating allele frequency distributions that were used to estimate the genetic information content of these loci, including genetic variability, inbreeding, individual and parent verification parameters. Disregarding MCMA26 monomorphic pattern, microsatellite loci showed moderate level of polymorphism, as such totally 102 alleles were detected with a mean number of 6 alleles per locus. The average expected heterozygosity was 0.72 (SD = 0.07) and the average Polymorphism Information Content (PIC) was 0.67 (SD = 0.08). Total value of inbreeding based on marker data was estimated as -0.02 so it indicates that inbreeding occurred less than would be expected at random. The overall probability of identity considering all twelve independent loci combined was 3.148E-13 meaning lower than 1 in 31 trillions. The results of this study indicate, despite the selective breeding and closed flock system over a number of generations, a relatively high level of heterozygosity still exists in the representative sheep flock. The high degree of multiallelism and the clear and simple codominant Mendelian inheritance of the set of microsatellites used provide a powerful system for the unique identification of Afshari sheep individuals for fingerprinting purposes and parentage testing.
PDF Abstract XML References Citation
How to cite this article
S. Qanbari, M.P. Eskandari Nasab, R. Osfoori and A. Hagh Nazari, 2007. Power of Microsatellite Markers for Analysis of Genetic Variation and Parentage Verification in Sheep. Pakistan Journal of Biological Sciences, 10: 1632-1638.
DOI: 10.3923/pjbs.2007.1632.1638
URL: https://scialert.net/abstract/?doi=pjbs.2007.1632.1638
DOI: 10.3923/pjbs.2007.1632.1638
URL: https://scialert.net/abstract/?doi=pjbs.2007.1632.1638
INTRODUCTION
When, the breeding and production work confined to only a closed flock (population) is likely to cause a significant restriction of the genetic variation and this is one of the basic factors determining the efficiency of the breeding work. Hence, a difficulty in managing of a selective breeding program is loss of genetic variability and increases in inbreeding as a result of the inadvertent mating of related individuals. The effects of inbreeding in sheep include moderate to high inbreeding depression of survival and growth traits (Lamberson et al., 1984; Gama and Smith, 1993; Wiener et al., 1992). Inbreeding also will result in a decrease in genetic variability, which will limit the potential for genetic gain from artificial selection (Falconer and Mackay, 1996). Therefore, evaluating the polymorphism of the genetic structure of a given breeding flock (population) is important, as the variation level is a basic factor determining the selection efficiency in successive generations. On the other hand, for every program of breeding and selection, it is desirable to rely on a precise assessment of parentage to assist, for example, in the elimination of undesirable recessive alleles, in early selection for future breeding, or to ensure accurate pedigrees and registration (Tomasco et al., 2002).
The use of microsatellite regions, segments of the nuclear genome composed of tandem repeats of short-sequence motifs, is well established and accepted as a method for the study of genetic information content of animal populations (Goldestein and Schlotterer, 2000). A large number of highly polymorphic microsatellites have been characterized and mapped in domestic animals, including sheep, cattle and other ruminants (de Gortari et al., 1997; Hayes et al., 1996; Jenkings et al., 1997), facilitating the use of these markers for detailed investigation of the genetic structure of a population. In the case of genetic variability, the analysis of allelic distribution at highly polymorphic microsatellite loci can be used to monitor the genetic structure of populations and to detect changes in the frequency of alleles due to breeding. That also is proven, the microsatellite approach can be applied to the creation of mating schemes aimed to increase genetic variability, reduce consanguinity and enhance fitness within flocks (Tomasco et al., 2002).
Microsatellites, also known as SSRs, are certainly the most efficient way of resolving issues of parentage verification and individual identity (deNise et al., 2004; Luikart et al., 1999). Microsatellites are typically codominant and multiallelic, with expected heterozygosity frequently greater than 0.7, allowing precise discrimination of even closely related individuals. Due to the specificity of the PCR assay and its high information content, it also allows the determination of identity between individuals based on formal estimates derived from allele frequencies.
In this study, the use of microsatellite markers in the evaluation of genetic information content provides statistics on the current status of genetic variation of Afshari sheep elite breeding flock, which has had a close mating system and experienced specific selection pressures over successive generations. Our objectives in the present study are: (1) to estimate allele frequencies and characterize these SSR loci for several parameters of intra-flock diversity including number of alleles, expected heterozygosity, polymorphism information content and inbreeding, also (2) to verify the exclusion probabilities and power of discrimination of this set of loci for individual fingerprinting in this breeding population.
MATERIALS AND METHODS
Blood samples and DNA extraction: A random sample consisting of one hundred animals were selected to analyze the DNA variability of the flock. Peripheral blood samples taken from fully pedigreed pure breed Afshari sheep belonging to the Research Farm of the University of Zanjan, Faculty of Agriculture, were used as material. DNA was extracted from 1ml whole blood samples by salting out method according to Miller et al. (1998) with minor modifications.
Microsatellite genotyping: Eighteen microsatellite markers distributed across the ovine genome were selected. The primer pairs were selected from Melbourne University sheep genome databases (Maddox et al., 2001) available on the website: http://rubens.its.unimelb.edu.au/~jillm/jill.htm. Microsatellite loci DYMS1, OarFCB304, BM8125, MAF65, OarJMP58, OarJMP29, are also belonging to the sheep biodiversity panel. Primer sequences, details of PCR conditions, corresponding chromosomal location, references and some other characteristics of the loci studied are described in Table 1. The PCR reactions were carried out in a thermocycler (i-Cycler, Biorad) with the following conditions: Each 15 μL PCR reaction contained 25 ng template DNA, 200 μM each of dNTP, 2.5 pmol each primer, 1X PCR standard reaction buffer and 1 unit of Biotherm Taq polymerase, that MgCl2 concentration was optimized for each microsatellite locus. After an initial denaturation at 94�C for 5 min, PCR products were loaded to 0.8 mm thick, 8% denatured polyacrylamide gel to electrophoresis in 0.5X TBE buffer with power 15 W (400 V, 35 mA) for 5 h. Allelic fragment sizes were determined using One-D-Scan software (SCAN ANALYTIC. CO) comparing to molecular weight standards.
Data analysis: Genotypes were assigned for each animal based on allele size data. On the basis of allele and genotypic frequencies, a likelihood ratio test (G2T) was conducted to test for deviations from Hardy-Weinberg equilibrium (Guo and Thompson, 1992). The most common measures of genetic diversity such as allelic diversity, heterozygosity and proportion of polymorphic loci were considered. The effective number of alleles (estimates the reciprocal of homozygosity) was calculated according to Hartl and Clark (1989). Observed (HO) and Nei unbiased expected heterozygosity (HE =1- Σp2i, where pi is the frequency of allele i) were estimated for all loci (Nei, 1978). These parameters were statistically analyzed using PopGene software package version 1.31 (Yeh et al., 1999). Polymorphism information content (PIC) (Botstein et al., 1980) values were estimated in order to assess the relevance of each locus for linkage analysis using following formula:
where pi and pj are the frequencies of the ith and jth alleles in the population, respectively and n is the number of alleles per locus.
In order to measure an individual’s inbreeding coefficient, detailed pedigree information is required. In recent years, the developments of molecular techniques offer the opportunity to determine accurate pedigrees in populations. Such approaches have the advantages that pedigree information is not required. The Inbreeding coefficient was calculated as the difference between observed and expected heterozygosity (Lukas and Donald, 2002).
where F is the coefficient of inbreeding, the deviation of the observed heterozygosity of an individual relative to the heterozygosity expected under random mating (Hardy-Weinberg equilibrium); HO is the observed frequency of heterozygous individuals and HE is the expected frequency of heterozygous in the population. When F is >0 it signifies more inbreeding than is expected at random and when F is <0 it indicates that inbreeding occurred less often than would be expected at random.
| Table 1: | Characteristics of Microsatellite loci screened for the analysis of intra-population variation, paternity and estimation molecular inbreeding metrics |
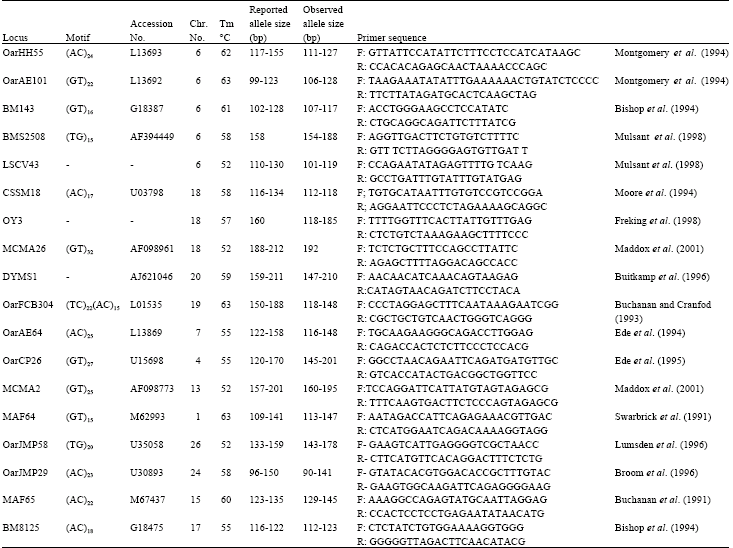 | |
We calculated probability of identity as individual differentiation parameter using the following formula:
(Paetkau et al., 1995), where pi and pj are the frequencies of the ith and jth alleles in the population. If I is the confusion probability, i.e., that two randomly chosen individuals from a sample have identical banding patterns, then discrimination power (D = 1-I) represents the probability that two randomly chosen individuals have different patterns and thus distinguishable from one another. Combined probability of genetic identity, CI =lll was estimated for the combined battery of studied loci.
The exclusion power (PE) for the parents when genomic DNA of both parents and one offspring was available was calculated using the formula of Jamieson and Taylor (1997):
After calculation of PE for each locus individually, a combined PE over n independent markers was calculated using the following formula: PE= 1- (1-P1) (1-P2) (1-P3) … (1-Pn) (Jamieson and Taylor, 1997).
Paternity exclusion probability (Q) (Weir, 1996), which corresponds to the power with which a locus excludes an erroneously assigned individual animal from being the parent of an offspring was estimated as;
where p is the population frequency of the ith or jth allele. The combined probability of paternity exclusion was calculated using the following formula:
RESULTS AND DISCUSSION
All eighteen markers subjected in this research were amplified in Afshari sheep. Microsatellite marker MCMA26 showed monomorphic pattern. As many population studies in the literature are based on polymorphic data only, it was necessary to exclude monomorphic data from our estimates for comparison with results of these studies (Bowcock et al., 1991; O’Brien et al., 1994; Kidd et al., 1991). Table 2 summarizes the statistics of genetic variation. Disregarding monomorphic locus, a total of 102 alleles were detected with the mean of 6 alleles per locus. Polymorphism was low at CSSM18, OarAE64 and BM8125 loci, each comprising 4 alleles and moderately high at other loci with up to 9 alleles. It has been shown that increasing the number of alleles at different loci increased the mean genetic diversity in population (Moioli et al., 2001). High mean number of observed alleles (allelic diversity) per locus displayed high genetic variation and indicates that the population is under mutation drift equilibrium. Despite the enormous increase in studies reporting genetic variation in sheep, little is known regarding variation at the DNA level within individual sheep population. For comparison, the mean allelic variability detected by genotyping of twenty four microsatellites loci in a captive and unmanaged population of St Kilda Soay sheep was reported as 5.42 (Overall et al., 2005). However, while there is no published literature on microsatellite variation in closed breeding flocks for comparison, the allelic variation at the studied loci was compared to those in previous reports available on sheep genome database (Maddox et al., 2001). On the basis of this comparison, allelic variability of all studied loci was declined dramatically. As shown in Table 1, the allele size range for some loci was not in coincidence with those reported in database, so they can be considered as new alleles for these loci. As a most common parameter for population variability an estimate of both observed (HO) and expected heterozygosity (HE) were estimated for each locus (Table 2). Microsatellite marker OarJMP58 having 9 alleles indicated the highest HE (85%). In contrary, the lowest HE was shown by CSSM18 locus as 0.61. Mean HE and mean Ho from data on seventeen polymorphic loci was estimated as 0.72 (SD = 0.07) and 0.74 (SD = 0.13) respectively. In the later study heterozygosity estimates computed across the 18 microsatellite loci for six closely related populations of Merino sheep breed ranged from 0.679 to 0.763. In similar studies, the high mean heterozygosity values have been attributed to low level of inbreeding, low selection pressure and large number of alleles present in a population (Arora and Bhatia, 2004). As shown in Table 2 total value of inbreeding based on marker data was estimated as -0.02 so it indicates that inbreeding occurred less than would be expected at random. In closed breeding flocks, success in controlling inbreeding over the long term will depend on our ability to limit genetic relationships between rams entering in mating. In the case of present study, observed level of heterozygosity and inbreeding for studied loci, could be explained as the result of controlled mating system over generations that have been implemented through breeding program aimed to maximize the retention of genetic variability and maintenance of lower inbreeding levels in the flock.
| Table 2: | Parameters of genetic information content of seventeen microsatellite loci estimated from the breeding flock of Afshari sheep |
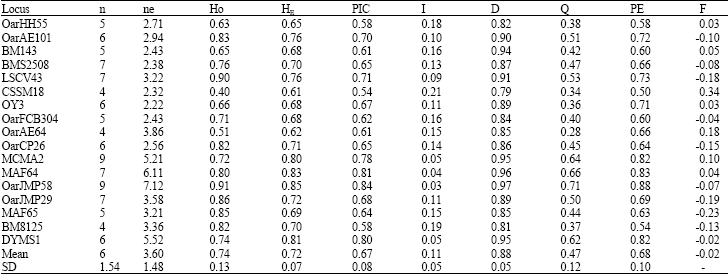 | |
Abbreviations are; Number of detected alleles (n), effective allele number, observed (Ho) and expected (HE) heterozygosity per locus, PIC value, probability of identity (I), discrimination power (D), Exclusion power (EPR), Probability of exclusion (PE) and inbreeding coefficient (F) | |
The PIC values at each marker were estimated in order to assess the relevance of each locus for linkage analysis (Table 2). Microsatellites markers having PIC values higher than 0.5 are considered highly informative (Botstein et al., 1980). The majority of the microsatellite loci studied would provide an excellent contribution to a genome scan with more than 65% of the meiosis expected to be informative in this population. These high estimates of PIC substantiated the suitability of used set of DNA markers to applications such as parentage control and linkage-mapping programs.
We also calculated the individual differentiation and parentage verification parameters such as I, D, Q and PE at each locus (Table 2). It is important to recognize that the formulas for estimating identity and exclusion probabilities assume random mating and non-random association among alleles and loci (i.e., Hardy-Weinberg proportions and no linkage disequilibrium). These assumptions are occasionally violated, especially when there are close relatives in the breeding population. These violations could cause an overestimation (bias) of the true probabilities and they are known to occasionally cause a bias of 1-3 orders of magnitude of the probability of identity (I) in wildlife populations (Waits et al., 2001). Other potential sources of error for individual and parentage verification are departures from Hardy-Weinberg proportions and occurrence of mutations as well. In the case of the present study, all loci significantly (p<0.01) deviate from Hardy-Weinberg proportions. Also mutation rates for many microsatellites are high enough (e.g., ~1.10-4) to occasionally cause errors in paternity exclusion. When loci are inherited independent of each other, the total values of aforementioned parameters are a sequential product of individual values of the loci. In such a way, theoretical cumulative values calculated by this set of markers may not reflect the actual values in a population due to the presence of linkage in some loci (chromosome 6 having five loci and 18 with two loci) and it becomes necessary to also recalculate them. Hence, cumulative values were estimated excluding of linked loci and including only the most effective polymorphic locus at each common chromosome. Consequently CI for a panel of twelve independent loci (OarJMP58, OarJMP29, OarCP26, MCMA2, CSSM18, MAF64, MAF65, BM8125, OarFCB304, DYMS1, LSCV043 and OarAE64) were estimated as 3.128E-13, meaning that the chance of finding two individuals with the same genotype in the population is almost null, hence D was close to 1. The exclusion probability (Q) indicates the probability of excluding a nonparent from paternity or maternity. The Q was moderate for all the microsatellites analyzed. It ranged from 0.38 (OarHH55) to 0.71 (OarJMP58), with an average of 0.47. The combined probability of paternity exclusion (QC) and combined exclusion power (CPE) for panel of independent loci were estimated as 1.75E-4 (99.98%) and 2.305E-7, respectively, indicating that these loci are appropriate to determine parentage in Afshari breeding flock beyond any reasonable doubt.
CONCLUSION
This research presents an initial step in investigation of variability at the DNA level within elite breeding flock of Afshari sheep. In the present study we have investigated the intra-population variation of the flock which has had a close mating system and experienced specific selection pressures over several successive years which, subsequently could have resulted in the reduction of genetic diversity due to an increase of inbreeding in the population. The results of this study indicate, despite the selective breeding and closed flock system over a number of generations, a relatively high level of heterozygosity still exists in the representative sheep flock. The Value of inbreeding level estimated based on marker data approved effectiveness of the mating scheme implemented over last generations. Also on the basis of the present study it can be concluded that the microsatellite markers can be used as an appropriate tools for individual identification, parentage verification and also to infer the levels of inbreeding directly from sampled individuals. The significance of this report is that it offers interesting perspectives for the incorporation of molecular genetic techniques to animal breeding in Iran. In addition the results of this study could provide basic molecular data for the research on germplasm characteristics of Afshari sheep.
ACKNOWLEDGMENTS
This study was conducted at the Animal Science Department of Research Institute of Physiology and Biotechnology. Our sincere thanks to H. Saeed Mohammadi for gathering and monitoring of pedigree data, M. Razavi Nia and H, Mehri for laboratory practices and Dr M. E. Fooladvand for helps in data analysis.
REFERENCES
- Bishop, M.D., S.M. Kappes, J.W. Keele, R.T. Stone and S.L.F. Sunden et al., 1994. A genetic linkage map for cattle. Genetics, 136: 619-639.
Direct Link - Botstein, D., R.L. White, M. Skolnick and R.W. Davis, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet., 32: 314-331.
PubMedDirect Link - Bowcock, A.M., J.M. Herbert and J.L. Mountain, 1991. Study of an additional 58 DNA markers in five human populations from four continents. Gene Geograph., 5: 151-173.
Direct Link - Broom, J.E., M.L. Tate and K.G. Dodds, 1996. Linkage mapping in sheep and deer identifies a conserved Pecora ruminant linkage group orthologous to two regions of HSA 16 and a portion of HSA7Q. Genomics, 33: 358-364.
Direct Link - Buchanan, F.C., P.A. Swarbrick and A.M. Crawford, 1992. Ovine dinucleotides repeat polymorphism at the MAF65 locus. Anim. Genet., 23: 85-85.
Direct Link - Buchanan, F.C. and A.M. Crawford, 1993. Ovine microsatellites at the OarFCB11, OarFCB128, OarFCB193, OarFCB266 and OarFCB304 loci. Anim. Genet., 24: 145-145.
PubMed - Buitkamp, J., P. Filmether, M.J. Stear and J.T. Epplen, 1996. Class I and class II major histocompatibility complex alleles are associated with fecal egg counts following natural, predominantly Ostertagia circumcincta infection. Parasit. Rev., 82: 693-696.
Direct Link - De Gortari, M.J., B.A. Freking, S.M. Kappes, K.A. Leymaster, R.T. Stone, C.W. Beattie and A.M. Crawford, 1997. Extensive genomic conservation of cattle microsatellite heterozygosity in sheep. Anim. Genet., 28: 274-290.
CrossRefDirect Link - DeNise, S., E. Johnston, J. Halverson, K. Marshall, D. Rosenfeld, S. McKenna, T. Sharp and J. Edwards, 2004. Power of exclusion for parentage verification and probability of match for identity in American kennel club breeds using 17 canine microsatellite markers. Anim. Genet., 35: 14-17.
Direct Link - Ede, A.J., C.A. Pierson, H.M. Henry and A.M. Crawford, 1994. Ovine microsatellites at the OarAE64, OarHH22, OarHH56, OarHH62 and OarVH4. Anim. Genet., 25: 51-52.
Direct Link - Ede, A.J., C.A. Pierson and A.M. Crawford, 1995. Ovine microsatellites at the OarCP9, OarCP16, OarCP20, OarCP21, OarCP23 and OarCP26 loci. Anim. Genet., 26: 129-130.
Direct Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Freking, B.A., J.W. Keele, C.W. Beattie, S.M. Kappes and T.P.L. Smith et al., 1998. Evaluation of the ovine callipyge locus: I. Relative chromosomal position and gene action. J. Anim. Sci., 76: 2062-2071.
Direct Link - Gama, L.T. and C. Smith, 1993. The role of inbreeding depression in livestock production systems. Livest. Prod Sci., 36: 203-211.
Direct Link - Guo, S.W. and E.A. Thompson, 1992. Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics, 48: 361-372.
PubMedDirect Link - Hayes, H., C. Le Chalony, G. Goubin, D. Mercier, E. Payen, C. Bignon and K. Kohno, 1996. Localization of ZNF164, ZNF146, GGTA1, SOX2, PRLR and EEF2 on homologous cattle, sheep and goat chromosomes by fluorescent in situ hybridization and comparison with t he human gene map. Cytogenet. Cell Genet., 72: 342-346.
Direct Link - Jamieson, A. and St. C.S. Taylor, 1997. Comparison of three probability formulae for parentage exclusion. Anim. Genet., 28: 397-400.
Direct Link - Jenkins, Z.A., H.M. Henry, S.M. Galloway, K.G. Dodds and G.W. Montgomery, 1997. Comparative linkage mapping of genes on sheep chromosome 3 provides evidence of chromosomal rearrangements in the evolution of bovidae. Cytogenet. Cell Genet., 78: 272-274.
Direct Link - Kidd, J.R., F.L. Black K.M. Weiss and K.K. Balazs Kidd, 1991. Studies of three Amerindian populations using nuclear DNA polymorphism's. Human Biol., 63: 775-794.
Direct Link - Luikart, G., M.P. Biju-Duval, O. Ertugrul, Y. Zagdsuren, C. Maudet and P. Taberlet, 1999. Power of 22 microsatellite markers in fluorescent multiplexes for parentage testing in goats (Capra hircus). Anim. Genet., 30: 431-438.
PubMedDirect Link - Keller, L.F. and D.M. Waller, 2002. Inbreeding effects in wild populations. Trends Ecol. Evol., 17: 230-241.
Direct Link - Lumsden, J.M., E.A. Lord and G.W. Montgomery, 1996. Characterization and linkage mapping of ten microsatellite markers derived from a sheep x hamster cell hybrid. Anim. Genet., 27: 203-206.
Direct Link - Maddox, J.F., K.P. Davies, A.M. Crawford, D.J. Hulme and E.P. Cribiu et al., 2001. An enhanced linkage map of the sheep genome comprising more than 1000 loci. Genome Res., 11: 1275-1289.
Direct Link - Miller, S.A., D.D. Dykes and H.F. Polesky, 1988. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res., 16: 1215-1215.
PubMedDirect Link - Moioli, B., A. Georgoudis, F. Napolitano, G. Catillo, E. Giubilei, C. Ligda and M. Hassanane, 2001. Genetic diversity between Italian, Greek and Egyptian buffalo populations. Livest. Prod. Sci., 70: 203-211.
Direct Link - Montgomery, G.W., E.A. Lord, J.M. Penty, K.G. Dodds and T.E. Broad et al., 1994. The booroola fecundity (FecB) gene maps to sheep chromosome 6. Genomics., 22: 148-153.
CrossRefPubMedDirect Link - Moore, S.S., K. Byrne, K.T. Berger, W. Barendse, F. McCarthy, J.E. Womack and D.J. Hetzel, 1994. Characterization of 65 bovine microsatellites. Mamm. Genome, 5: 84-90.
Direct Link - Nei, M., 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590.
PubMedDirect Link - O`Brien, E., A.R. Rogers, J. Beesley and L.B. Jorde, 1994. Genetic structure of the utah mormons: Comparison of results based on RFLPs, blood groups, migration matrices, isonymy and pedigrees. Human Biol., 66: 743-759.
Direct Link - Overall, A.D.J., K.A. Byrne, J.G. Pilkington and J.M. Pemberton, 2005. Heterozygosity, inbreeding and neonatal traits in Soay sheep on St Kilda. Mol. Ecol., 14: 3383-3393.
Direct Link - Paetkau, D., W. Calvert, I. Stirling and C. Strobeck, 1995. Microsatellite analysis of population structure in Canadian polar bears. Mol. Ecol., 4: 347-354.
CrossRefDirect Link - Swarbrick, P.A., F.C. Buchanan and A.M. Crawford, 1991. Ovine dinucleotide repeat polymorphism at the MAF64 locus. Anim. Genet., 22: 375-376.
Direct Link - Tomasco, I., G. Wlasiuk and E.P. Lessa, 2002. Evaluation of polymorphism in ten microsatellite loci in Uruguayan sheep flocks. Gene. Mol. Biol., 25: 37-41.
Direct Link - Waits, L.P., G. Luikart and P. Taberlet, 2001. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol., 10: 249-256.
Direct Link