
Babak Chaloushi
Department of Biotechnology, Faculty for Agronomical and Animal Sciences, Tehran University, Karaj, Iran
Reza Zarghami
Agricultural Biotechnology Research Institute of Iran, Karaj, Iran
Cyrus Abd-Mishani
Department of Biotechnology, Faculty for Agronomical and Animal Sciences, Tehran University, Karaj, Iran
Mansour Omidi
Department of Biotechnology, Faculty for Agronomical and Animal Sciences, Tehran University, Karaj, Iran
Yusif M. Agayev
Agricultural Biotechnology Research Institute of Iran, Karaj, Iran
Babak Pakdaman Sardood
Agricultural Biotechnology Research Institute of Iran, Karaj, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 10 | Page No.: 1625-1631







ABSTRACT
Saffron (Crocus sativus L.) is one of valuable and native plants in the land of Iran. By this investigation the best hormonal compositions for callus production from protoplast and for plantlet regeneration from callus were determined. To isolate protoplasts, the embryogenic calli were used. The embryogenic calli were immersed in enzymatic solution to degrade the cell walls. The treated mixture was filtered and then centrifuged at 100 g for 3-5 min and the resulted pellet was rinsed. After one step of washing and another step of centrifugation, the protoplasts were gently mixed with sterile sodium alginate solution and added to MS broth consisting 1% calcium chloride and 0.3 M manitol to form calcium alginates granules. The protoplast-containing granules were exposed to MS broth including 0.3 M manitol and various treatments of two kinds of auxins (2, 4-D and NAA) and three kinds of cytokinins (2ip, Kin, BAP), respectively in four rates of 0.1, 0.2, 0.5, 1.0 mg L-1 for auxins and in three rates of 0.1, 0.2 and 0.5 mg L-1 for cytokinins, incubated in dark at 22 ± 2 °C for a period of 30 days. Out of all the treatment of 2, 4-D (1.0 mg L-1) and Kin (0.2 mg L-1) was the best in callus induction. In order to regenerate plantlets, the resulted calli were transferred to MS broth amended with different rates of ABA (0.0, 0.5, 1.0, 1.5, 2.0 and 2.5 mg L-1) so that they could pass the steps of embryonic maturation. The mature embryos were transferred to MS media with different rates of GA3 (0.0, 5.0, 15.0, 20.0, 25.0 and 30.0 mg L-1) to initiate germination. The germinated embryos were then placed in solid MS media with various rates of NAA and 2, 4-D auxins (0.0, 0.5, 1.0 and 2.0 mg L-1) and different levels of BAP and Kin cytokinins (0.0, 0.5, 1.0 mg L-1). Results from statistical analyses indicated the treatment of NAA and BAP (each 1 mg L-1) as the best hormonal treatment for the plantlet regeneration from the domestic saffron calli.
PDF Abstract XML References Citation
How to cite this article
Babak Chaloushi, Reza Zarghami, Cyrus Abd-Mishani, Mansour Omidi, Yusif M. Agayev and Babak Pakdaman Sardood, 2007. Effects of Different Hormonal Treatments on the Callus Production and Plantlet Regeneration in Saffron (Crocus sativus L.). Pakistan Journal of Biological Sciences, 10: 1625-1631.
DOI: 10.3923/pjbs.2007.1625.1631
URL: https://scialert.net/abstract/?doi=pjbs.2007.1625.1631
DOI: 10.3923/pjbs.2007.1625.1631
URL: https://scialert.net/abstract/?doi=pjbs.2007.1625.1631
INTRODUCTION
Saffron (Crocus sativus L.) as the most expensive agronomical and pharmacological crop in the world is of particular position among agricultural crops, native in the Iran and cultivated in most eastern and central regions of the country (Abrishami, 1987). Saffron is a perennial, herbaceous plant belonged to the family Iridaceae and asexually propagated through it’s corms. Till now, nine species of saffron has been reported from Iran (Abrishami, 1987; Kafi, 2000). Saffron is applicable in drug and food industries and has attracted attentions because of its biologically profitable impacts especially as a source of natural anti-carcinogenic compounds. To determine the quantities and qualities of saffron chemical ingredients, there have been performed many experiments indicating the most effective active ingredients of saffron as water-soluble carotenoid pigments, a bitter-tasting glycoside picrocrocin, aromatic volatile compounds mainly safranal and substances like carbohydrates, pectins, pentosanes, minerals, vitamins (B2) and fatty acids (palmitic acid, actearic acid and linoleic acid) (Radjabian, 2000). Because of triploidy, sterility, slow rate of growth and susceptibility to injuries, the prevalent methods for breeding are not mainly applied with saffron, therefore, establishment and development of a system for in vitro culture and propagation of saffron is the first step to approach to breeding goals, so that tissue-culture is now a daily regarded as an appropriate and useful method for genetic improvement of saffron and production of new varieties.
Briefly, the advantages of saffron tissue-culture and micropropagation are as: fast and large-scale propagation of new and improved varieties, or of selected ecotypes to provide healthy and intact corms; regeneration and propagation of new genotypes through application of somaclonal diversity; production of pathogen-free cormsand of healthy plants through generation of corms and somatic embryogenesis, protoplast culture, stamen culture, pollen culture, somatic hybridization between domestic saffron and it’s wild species and relatives, out of them, protoplast culture can be considered as one of the tissue-culture methods usable in saffron genetic breeding and the improvement of it’s properties (Bokelmann and Roest, 1983; Draget et al., 1988). The importance of protoplast culture technology in the studies on protoplasts conjugation, absorption of exterior DNA through plasmids and bacteria, callus production and plantlet regeneration, studies on the biosynthesis and reconstruction of cell walls and artificial seeds production is not ignorable. Taking advantage of MS culture medium and hormones 2, 4-D, Kin and NAA, Ding et al. (1979) could induce callus formation on the corm fragments and finally regenerate plantlets out from the calli for the first time Ilahi, Sano, Isa, Ogasawara, Ebrahimzadeh, Karamian, Kobayashi achieved success through production of strigma-like structures, calli and plantlets through cultivation of different fragments from saffron plants (Ebrahimzadeh et al., 2000a; Ilahi et al., 1987; Isa and Ogasawara, 1988). With an eye to the importance of saffron and the previously performed investigations, the present research was carried out to determine the best hormonal composition of the medium for production of calli from protoplasts and for regeneration of plantlets from the resulted calli.
MATERIALS AND METHODS
To isolate protoplasts, the embryogenic calli resulted from saffron corm mersitem cultures were applied. The embryogenic calli were immersed in an enzymatic solution including pectolyase Y-2-3 (0.1% W/V), cellulose R-10 (1% W/V), driselase (1% W/V), 4-2-N-morpholino ethanesulfonic acid (MES, 0.1% W/V) and manitol (0.3 M) prepared in MS medium and shaked to allow cell wall digestion.
The treated mixture was filtered through the filters with the pore size of 43-50 μm and then centrifuged at 100 g for 3-5 min. The resulted pellet was washed with washing solution (0.1% MES in 0.3 M manitol) and re-centrifuged.
After a further step of washing and centrifugation, the prepared protoplasts were studied from the stand point of their viability and using Evan’s blue stain. To cultivate protoplasts, the method of fixation in calcium alginate granules was followed and the protoplasts in a given density were mixed with the sterile sodium alginate solution (2% in 0.3 M manitol). Using a micropipette with a sterile tip, sodium alginate solution together with protoplasts was added drop by drop into the MS broth containing 1% calcium chloride and 0.3 M manitol, while the latter was regularly and gently stirred by a sterile magnet. Inside this solution, a granule or a bead of calcium alginate was instantly developed from each of the added drops. The granules were kept stationary in the solution for 15-20 min to allow their complete gelatinization and then washed twice with MS broth containing 0.3 M manitol. The protoplast-containing granules were exposed to MS broth including 0.3 M manitol and various treatments of two kinds of auxins (2, 4-D and NAA) and three kinds of cytokinins (2ip, Kin, BAP) respectively in four rates of 0.1, 0.2, 0.5, 1.0 mgL-1 for auxins and in three rates of 0.1, 0.2 and 0.5 mgL-1 for cytokinins, incubated in dark at 22±2°C, so that those were subcultured through replacement of the old and exhausted culture media by fresh and hypotonic culture media once per 7-10 days. A month after the formation of cellular masses and with the observation of the masses of microcalli developed from the granules, they were transferred to solid MS media with the same types and the same concentrations to accelerate callus formation.
To regenerate the plantlets, the following steps were attempted. At first, the resulted calli were transferred to solid MS culture media containing different rates of ABA (0.0, 0.5, 1.0, 1.5, 2.0 and 2.5 mg L-1) to attain somatic maturation. For growth and germination, the somatically mature embryos were then transferred to the MS medium supplemented with different amounts of GA3 (0, 5, 10, 15, 20, 25 and 30 mg L-1). To initiate plantlet development, the germinated somatic embryos were placed in the solid MS culture media synchronously amended with auxins, 2, 4-D and NAA (0, 0.5, 1 and 2 mg L-1) and the cytokinins, Kin and BAP (0, 0.5 and 1 mg L-1). The cultures were incubated at 20±2°C and under photoperiodic conditions of 16 h illumination and 8 h darkness. The cultures were subcultured in regular intervals of 21-28 days for the period of 3-4 months.
To determine the best hormonal treatment, the statistical method of none-parametric analysis of data was applied and data were tested by Kruskal-Wallis test and taking advantage of SPSS software.
RESULTS AND DISCUSSION
Isolation of protoplasts: The most abundant number of protoplasts were obtained when embryogenic calli were enzymatically digested 7-10 days after subculturings in fresh media (Fig. 1).
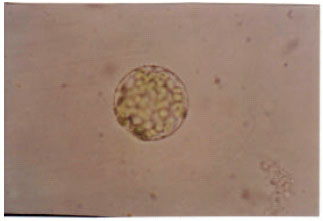 | |
| Fig. 1: | A protoplast with dense cytoplasmic contents isolated from embryogenic callus |
This fact indicates that at the first days of subculturing in fresh media, cells are of more growth activities that is because of the provision of the compounds required for more cellular growth and division and consequently, more protoplasts of more capabilities for growth and multiplication are generated. In deed, the quality and quantity of the isolated protoplasts depends on the features of the formed calli and the length of subculturing period. Microscopy of the calli treated with the enzymatic solution carried out in the intervals of 20-30 min revealed the occurrence of cell wall degradation within 2.5-3 h. These observations are in agreement with those by Isa and Ogasawara (1990), however somewhat different from the results got by Karamian (2000), who recognized a period of 1.5-2 h as an appropriate time course for cell wall degradation. This difference may be originated from the type of micro-samples, discrepancies in cell wall structure, enzyme concentrations and environmental situations (like temperature, illumination and so on). Also, in all cases scintillations during enzymatic treatments led to the disintegration of plasma membranes of the isolated protoplasts prior to their death. However, division of this phase to two stages with and without shaking results in better results.
Protoplast culture and callus induction: Through observations made with granules under an inversed microscope (Nikon, Eclipse TE 300), it was demonstrated that in most cases regeneration of protoplast cell walls and the primary division of the regenerated cells occurred within 3-4 days. However, only with some of these treatments, they could continue later divisions and form cellular aggregates (Fig. 2).
In addition, compared with free control protoplasts, the protoplasts trapped in calcium alginate gel yielded better results from the stand point of cell division and aggregation (Table 1).
| Table 1: | Effect of Ca-alginate granules on the growth of saffron protoplasts |
A: Protoplast culture in a liquid medium without Ca-alginate mediated stabilization; B: Protoplast culture in a liquid medium with Ca-alginate mediated stabilization; CD: Cellular division; CM: Cellular mass; ICM: Increased cellular mass; MF: Microcallus formation | |
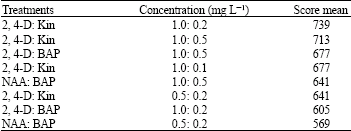
| Table 2: | Scores of different hormonal treatments callus-inducive effects |
 | |
| Table 3: | Scores of different rates of ABA based on their effects on embryogenic maturation |
 | |
Accomplishments through the application of calcium alginate as a base for protoplast culture have been subject of several reports in literature (Karamian, 2000; Brodelious and Nilsson, 1980; Draget et al., 1988). It seems that sensitive and brittle cells like protoplasts of plant origins are well protected following this approach, because this substance is not toxic and of deleterious effects against protoplasts and its gelatinization is independent on temperature. Also, calcium alginate provides an even environment around plant protoplasts and thereby helps them considerably in nutrient absorption (Karamian, 2000; Draget et al., 1988; Scheurich et al., 1999).
Out of all the treatments, the highest level of callus induction was attained taking advantage of 2, 4-D and Kin with the respective concentrations of 1 and 0.2 mg L-1, followed by the treatments with 2, 4-D + BAP and also NAA + BAP (df = 71, p>99%; Table 2). Furthermore, although the growth rate of calli was high with the hormonal treatments of 2, 4-D and Kin, however, calli induced by these treatments were a little darker unless they were subcultured in temporal intervals less than a month.
Regeneration of plantlets: Significant differences were found among the effects of all ABA rates tested (df = 5; p>99%). Comparison of the scores (Table 3) revealed the more effective role of 1 mg ABA L-1 in the maturation of the embryos and in the embryonic stages (Fig. 3).
| Table 4: | Scores of different GA3 rates |
 | |
| Table 5: | Scores of different hormonal treatments based on their effects on plantlet regeneration |
 | |
| Table 6: | Scores of different rates of NAA-BAP hormones based on their effects on plantlet regeneration |
 | |
It appears that ABA acts as an effective inducer of the expression of the genes, involved in maturation and inhibits embryonic pre-germination (Iida et al., 1992).
Transportation of the mature embryos to the culture media with higher concentrations of GA3 promoted their growth and germination. Statistically significant differences were found among the impacts imposed by the various concentrations of the hormone GA3 (df = 5, p>99%).
Comparison of the scores (Table 4) showed that the treatment with 25 mg GA3 L-1 had the best impression on the growth and germination of the matured embryos (Fig. 4). These findings are in accordance with the results from studies made by Karamian (2000).
Additionally, different auxin-cytokinin combinations exerted significantly different effects on the regeneration of saffron plantlets (df = 3; p>99%), so that, in accordance with results obtained by Ebrahimzadeh and coworkers (2000), the treatments with NAA-BAP was of more effects on the regeneration of the germinated embryos when scores from various hormonal combinations were compared (Table 5).
Out of all the concentrations of NAA and BAP applied in paired combinations, the pair composed of 1 mg NAA L-1 and 1 mg BAP L-1 was the most meritorious hormonal combination to regenerate saffron plantlets (df = 11, p>99%; Table 6).
One of the crucial points in the successful regeneration of the saffron plantlets is the incubation of the cultures at the temperatures of 20±2°C or less and under suitable photoperiodic conditions. Incubation of the calli at even lower temperatures significantly increased plantlet regeneration rate (Fig. 5).
Isa and Ogasawara (1988) had previously reported the positive effect of lower temperatures (15-20°C) on the regeneration of plantlets from embryogenic and non-embryogenic calli, from domestic saffron (Isa and Ogasawara, 1988; Isa et al., 1990). With an eye to the results from the studies on the protoplast cultures of domestic saffron, it can be concluded that various factors as temperature, illumination, method of cultivation and concentration of the growth regulators impose their effects on the growth and division of protoplasts and in next steps, on the growth and maturation of somatic embryos and finally on the formation of plantlets from the somatic embryos obtained from protoplasts (Bokelmann and Roest, 1983; Ebrahimzadeh et al., 2000b; Isa et al., 1990).
Therefore, the isolation of the protoplasts from embryogenic cultures can considerably help the development of synthetic seed production technology.
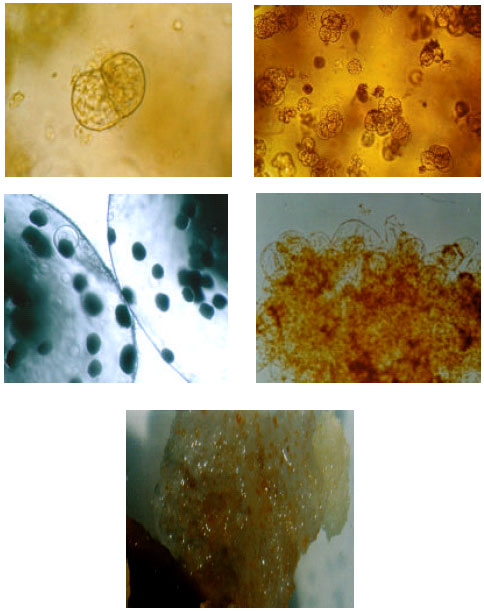 | |
| Fig. 2: | Stages of callus development from the cultivated protoplast of saffron: (A) Primary division of the cell wall- developed protoplast; (B) Development of cellular masses; (C) Increased cellular masses in Ca-alginate granules; (D) microcallus and (E) callus |
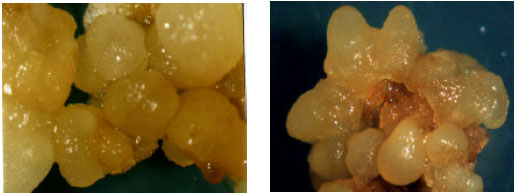 | |
| Fig. 3: | Stages of embryogenesis: (A) spherical (globular) stage of embryogenic callus; (B) Horn-shaped stage of embryogenic callus |
 | |
| Fig. 4: | Formation and development of leaf-like aerial parts from a mature embryo |
 | |
| Fig. 5: | In vitro development of (A) aerial parts, (B) green chlorophyll-containing leaf-like aerial part, (C) corm and (D) a mature corm formed and developed in vitro |
ACKNOWLEDGMENTS
Authors thank Mr. Dr. Hassan Rahnama, Mr. Dr. Dariush Davoudi and Mrs. Dr. Maryam J. Kermani at the Agricultural Biotechnology Research Institute of Iran, Karaj, who kindly helped us by their scientifically valuable guidance and encouragements.
REFERENCES
- Ebrahimzadeh, H., R. Karamian and M.R. Nori-Daloii, 2000. Somatic embryogenesis ad regeneration of plantlet in saffron, Crocus sativus L. J. Sci. I. R. Iran, 11: 169-173.
Direct Link - Ebrahimzadeh, H., R. Karamian and M.R. Nori-Daloii, 2000. Shoot regeneration from saffron protoplasts immobilized in Ca-alginate beads. J. Sci. I. R. Iran, 11: 69-72.
Direct Link - Iida, Y., K. Watabe, H. Kamada and H. Harada, 1992. Effects of abscisic acid on the induction of desiccation tolerance in carrot somatic embryos. Plant, 140: 356-360.
Direct Link - Isa, T., T. Ogasawara and H. Kaneko, 1990. Regeneration of saffron protoplasts immobilized in Ca-alginate beads. Jap. J. Breed., 40: 153-157.
Direct Link