
L. Naraghi
Plant Pests and Diseases Research Institute, Yaman Street, Chamran Free Way, P.O. Box 1454, Tehran 19395, Iran
H. Zareh-Maivan
Tarbiat Modarres University, Tehran, Iran
A. Heydari
Plant Pests and Diseases Research Institute, Yaman Street, Chamran Free Way, P.O. Box 1454, Tehran 19395, Iran
H. Afshari-Azad
Plant Pests and Diseases Research Institute, Yaman Street, Chamran Free Way, P.O. Box 1454, Tehran 19395, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 10 | Page No.: 1596-1603







ABSTRACT
In this study, combinations of spores of a thermophilic fungus (Talaromyces flavus), Vesicular Arbuscular Mycorrhiza (VAM) and microsclerotia of Verticillium dahliae under various temperature treatment (31-38°C) in triplicate trial, were investigated and results were compared with those of non-treated controls. Five cotton seeds were planted in each pot containing 3 kg of pasteurized soil. In each pot, combinations of 4x103 VAM spores, 2.5x10 9 spores of thermophilic fungus (T. flavus ) and 5x105 microsclerotia of V. dahliae were added. Symptoms of Verticillium wilt were observed after 45 days. Index of disease severity was measured. Results indicated that pre-heating of microsclerotia at 31 and 35°C for 10 and 14 h, respectively, caused a 15% reduction in leaf infection index. Presence of VAM and thermophilic fungus (T. flavus) spores caused 23 and 50% reductions in the disease development , respectively. Concurrent presence of mycorrhiza and thermophilic fungus spores caused a 10-20% reduction in disease development. These findings provide a promising approach to the control of Verticillium wilt of cotton. However, heat treatment of soil may prove difficult. Further studies in this regard are required and useful agricultural practices such as seasonal heating may be applied in the cotton fields.
PDF Abstract XML References Citation
How to cite this article
L. Naraghi, H. Zareh-Maivan, A. Heydari and H. Afshari-Azad, 2007. Investigation of the Effect of Heating, Vesicular Arbuscular Mycorrhiza and Thermophilic Fungus on Cotton Wilt Disease. Pakistan Journal of Biological Sciences, 10: 1596-1603.
DOI: 10.3923/pjbs.2007.1596.1603
URL: https://scialert.net/abstract/?doi=pjbs.2007.1596.1603
DOI: 10.3923/pjbs.2007.1596.1603
URL: https://scialert.net/abstract/?doi=pjbs.2007.1596.1603
INTRODUCTION
Plant diseases are mostly controlled by chemical pesticides and in some cases by cultural practices. However, the widespread use of chemicals in agriculture has been a subject of public concern and scrutiny due to the potential harmful effects on the environment, their undesirable effects on non-target organisms and possible carcinogenicity of some chemicals (Cook and Baker, 1983). Other problems include development of resistant races of pathogens, a gradual elimination and phasing out of some available pesticides and the reluctance of some chemical companies to develop and test new chemicals due to the problems with registration process and cost. The need for the development of non-chemical alternative methods to control plant diseases is therefore clear (Cook and Baker, 1983). Biological control of plant diseases has been considered a viable alternative method to manage plant diseases (Atkinson et al., 1994; Borowicz, 2001). Biocontrol is environmentally safe and in some cases is the only option available to protect plants against pathogens (Atkinson et al., 1994).
Cotton is an important cash crop which is being cultivated in many countries. Like other crop plants, cotton is also susceptible to several plant pathogenic agents including fungi. Verticillium wilt and seedling damping-off are considered the most important diseases of cotton in Iran (Hamdollahzadeh, 1993; Mansoori and Hamdollahzadeh, 1995). For controlling Verticillium wilt of cotton cultural practices and the use of resistant varieties are the most common strategies which are not either available or effective. Biological methods using fungal and bacterial antagonists in recent years have been applied to control cotton diseases (Heydari et al., 1997; Heydari and Misaghi, 1998; Heydari and Misaghi, 2003; Dutta, 1981; Spinks and Rowe, 1989; Aziz, et al., 1997).
Talaromyces flavus is a fungal antagonist that is concerned to thermophilic fungi groups and has been used in biological control of Verticillium wilt of cotton (Marois et al.,1984; Tjamos and Fravel, 1995; Naraghi et al., 2004). The fungus produces and secrets some metabolites which are proposed to be responsible for its antagonistic activity. Glucose oxidase is one of these metabolites that is taught to play an important role in fungal antagonism.
In an experiment, the glucose oxidase enzyme encoding gene was isolated from a T. flavus isolate and was inserted to Aspergillus niger a fungus with very similar amino acid sequence. The transformed A. niger showed in vitro antagonistic activity against V. dahliae by inhibiting its growth when its culture filtrate was tested on V. dahliae indicating the role of glucose oxidase enzyme in the antagonism (Murray et al., 1997). In addition to glucose oxidase other secreted metabolites are possibly involved in T. flavus antagonistic activity against V. dahliae (Murray et al., 1997). Research about the effect of heat treatment on the antagonistic effects and the enzymatic activity of thermophilic fungi have shown that pre-heating of microsclerotia under different heating programs using suitable times and temperature, has caused a significant increase in the antagonistic activity of thermophilic fungal against V. dahliae (Tjamos and Fravel, 1995).
Studies on possible function of VAM (Vesicular arbuscular mycorrhiza) for protection against plant pathogens, began in 1970s. It is apparent from the investigations on VAM-plant pathogen interactions that VAM can usually deter or reduce the severity of disease caused by soil-borne plant pathogens (Dehne, 1982). In a related study, Borowicz found that VAM can affect and influence plant-pathogen interaction (Borowicz, 2001). In other studies, the effects of VAM on shoot apical growth of micro propagated apple and development of micro propagated plum shoots (Fortuna et al., 1996 and 1998) and development of Verticillium and Fusarium wilt of alfalfa (Hwang et al., 1992) were investigated. Research about the effect of heating on VAM showed that pre-heating of the soil caused the growth-enhancement of VAM by Composted Grape Pomace ( CGP) or its extract. The effect could have been due to providing a source of P to CGP that overcomes the P-limiting conditions, and to the mycorrhizal fungus P uptake enhancement (Linderman and Davis, 2001). Gavito and Olsson (2003) hypothesized that soil temperature would have a higher impact on VAM development and nutrient uptake by the plant. VAM development and function is likely more influenced by the temperature component of climate change than by its [CO2] component.
For further investigation of interactions among the above-mentioned factors this study was conducted. In this study the effect of a combination of thermophillic fungus, mycorhizza and heating on the incidence and development of cotton Verticillium wilt disease was investigated.
MATERIALS AND METHODS
Isolation of Verticillium dahliae from soil: During 1998-99, field soil samples (15% clay, 33% silt and 52% sand; pH, 7.8) were collected from cotton field of Karkandeh Research Station in Golestan province of Iran and were carried to the laboratory using plastic bags. Samples were air-dried, homogenized using a revolving jar mill and stored at 4°C. For isolation of V. dahliae from soil the wet sieving procedure (Huisman and Ashworth, 1974) was carried out.
Pathogenicity test: Three fungal microsclerotia were randomly selected and cultured on C Zapeck solution agar medium. After 7 to 10 days, a fungal suspension with the concentration of 106 spores per mL was prepared for inoculation of cotton seedlings (Sahel cultivar). For inoculation, five cotton seedlings at 4 to 8 leaves stages were selected and were inoculated using stem puncture method. Control seedlings were treated only with distilled water. Disease incidence was evaluated five to six weeks after inoculation (Hamdollahzadeh, 1993). Fungal population in the soil was also evaluated according to procedure described by Booth (1970). For disease evaluation, infection degree was measured according to a 0-4 scale as follows: 0: No symptoms, 1: slight wilt and no defoliation, 2: perceptible wilt and defoliation, 3: wilt and defoliation more than 50%, 4: complete wilt and defoliation.
Preparation of the fungal inoculum for green house experiment: Preparation of V. dahliae microsclerotial suspension was carried out according to the procedure described by Tjamos and Fravel (1995). The slurry containing the microsclerotia was spread uniformly in a thin layer in 9 cm diameter glass Petri plate using approximately one plate for each original flask. The slurry was dried in a laminar air flow hood for 1, 2 h. Plates were then covered and transferred to a 30°C incubator for 30 h to destroy remaining hyphae and conidia. Microsclerotia from each plate were resuspended in 100 mL of SDW and macerated twice for 1 min in an Omnimixer at full speed. Suspension concerned to every plate was allowed to settle for 30 min and the top layer discarded and bottom layer was suspended in 3 mL of SDW in a 1.6 cm widex15 cm height test tube. Tubes were placed in water bath at temperatures corresponding to those found at 30 cm soil depth during solarization (Katan, 1987; Katan and De Vay, 1991). Regime 1 consisted of 31°C for 10 h followed by 35°C for 14 h. Regime 2 consisted of 33°C for 10 h followed by 36°C for 14 h. Regime 3 consisted of 35°C for 10 h followed by 38°C for 14 h. Nontreated microsclerotia were held under ambient conditions in water. Microsclerotia were heated for 3 days in a given regime (Tjamos and Fravel, 1995).
Pot soil inoculation was carried out as follows: Two hundred microsclerots were used for each g of pot soil. One milliliter of suspension containing heated microsclerotia was poured in a 2 cm diameter petri plate and the number of microsclerotia was determined using stereo dissecting microscope. This procedure was repeated for suspension containing non-heated microsclerotia as well.
Isolation of thermophilic fungus from soil: Cotton field soil collected from Karkandeh Research Station in Golestan province was used for isolation of thermophilic fungus (Talaromyces flavus) (Cooney and Emerson, 1964) Selective medium (TF medium) was adopted for isolation of the fungus from soil. This medium contained 1 L distilled water, 39 g Potato-Dextrose Agar (PDA), 2.0 mL of a 50% solution of lactic acid, 100 mg streptomycin sulfate, 50 mg chlorotetracycline HCl, 50 mg chloramphenicol, 4 mg pimaricin (2.5% sterile water suspension), 30 mg nystatine (Mycostatin, 4960 units mg-1) and 0.5 g oxgall (Bile, bovine). Lactic acid and the antimicrobial agents were added as aqueous solutions to autoclaved PDA at about 50°C. The medium was poured into petri dishes (18 mL per dish). One milliliter of aliquots was removed from 10-2 and 10-3 dilutions ( soil in water) during agitation with a magnetic stirrer and spread on the TF medium ( five plates per replication). Plates were incubated in the dark at 30°C for 7-8 days. T. flavus was detected and identified on the TF medium by its bright yellow colonies 10 days after incubation. Using the above-described procedure, propagules of T. flavus were isolated from soil (Marois et al., 1984).
Preparation of T. flavus inoculum for pot soil: Preparation of T. flavus inoculum was carried out according to the procedure described by Naraghi (2004).
T. flavus isolated from soil was cultured in 1.6 cm widex15 cm height test tubes contained TF medium. After 5 days, 20 mL SDW was poured into these tubes and mixed throughly. The content of each tube was then transferred into 50 cm widex80 cm-tall sterile plastic bag containing 250 g of wheat straws. Plastic bags were incubated at 30°C for 30 days. The contents of each plastic bag was evacuated and grinded by blender after covering the surface of wheat straws with T. flavus hyphae completely. Then, the number of ascospores of T. flavus in each g of grinded wheat straw was determined using a hemocytometer. In this procedure, 10 g ginded wheat straw was suspended in 10 mL SDW and the number of ascospores of T. flavus in 1 g of grinded wheat straw was determined by counting ascospores in one mL of this prepared suspension. For each g of soil, 0.52 g grinded wheat straw was used which contained 107 ascospores (Naraghi et al., 2004).
Isolation of VAM spores from soil: Cotton field soil was collected from Karkandeh Research Station in Golestan province for isolation of VAM spores. The procedure was similar to those of nematode isolation described by Jenkins and Taylor (1967).
Inoculation of soil with VAM spores: For inoculation of soil with VAM spores, field soil containing these spores was used. The amount of field soil added to each pot was adjusted so that each pot contained 4x103 VAM spores (Run-Jin, 1995). According to this procedure, 2.2 kg of infested field soil containing approximately 4x103 VAM spores was poured in each pot and the rest was filled with pasteurized soil.
Study of the antagonistic effects of thermophilic fungus, VAM association and heating on pathogenicity of Verticillium dahliae in the greenhouse. The experiment for the study of the effect of thermophilic fungus, mycorrhiza and heathing on the pathogenicity of V. dahliae was carried out as follows:
A completely randomized experiment with 20 treatments each with 3 replications were conducted in greenhouse conditions. Each replication consisted of a 7 cm diameter and 20 cm height plastic pot containing 2.5 kg sterilized soil and 3 seeds of Sahel cotton cultivar. In each pot, different combinations of VAM spores, spores of thermophilic fungus and microsclerotia of V. dahliae were added according to the experimental design. Leaf infection index (L.I) was determined according to the procedure described by Iakutkin and Popov (1979) as follows:
| L.I. | = | Leaf Infection Index |
| Infection Degree 1 | = | chlorosis and necrosis of 25% leaf surface |
| Infection Degree 2 | = | chlorosis and necrosis of 50% leaf surface |
| Infection Degree 3 | = | chlorosis and necrosis of 75% leaf surface |
| Infection Degree 4 | = | chlorosis and necrosis of more than 75% of leaf surface as well as defoliation |
| n | = | The number of leaf concerned to every infection degree |
| N | = | The number of all leaves of a plant |
Leaf Infection Index, the number and surface area of leaves and plants height were determined at 15 days intervals during three months after sowing and dry weight of plants was determined at the end of vegetative stage (9 months after sowing).
RESULTS
Verticillium wilt disease symptoms appeared 30 to 75 days after sowing. There was no significant differences in leaf infection index among different treatments before this (Table 4). Significant differences were observed among different treatments in the number of leaves 60 and 75 days after sowing (Table 3). The differences in leaf surface area and plant height were also significant among treatments from 15 to 45 days after sowing (Table 1 and Table 2). The alteration in Leaf Infection Index (LII) during 3 months after sowing, showed that L.I.I decreased from 45 to 75 days after sowing (Table 4). The significant increase in leaf surface area was observed among treatments contained both VAM spores and microsclerotia heated under program 3 compared to other treatments 15 days after sowing (Table 1). The increase in leaf surface area in treatments contained heated or non-heated microsclerotia combined with VAM and thermophilic fungus spores was significant compared to treatments without VAM spores 30 days after sowing (Table 1). However, no significant differences were observed in leaf surface area in different treatments 45 days after sowing. (Table 1). For seedling height, the significant increase were observed in treatments contained thermophilic fungi and VAM spores compared to those of controls (without VAM spores) 15 days after sowing (Table 2). Heating of V. dahliae microsclerotia also affected disease development and consequently the growth characteristics of cotton plants. Significant increases were observed in plant height in treatments contained both VAM spores and heated microsclerotia compared to those of non- heated 45 days after sowing (Table 2).
There was significant increase in leaf number in treatments contained microsclerotia heated under program 2 individually or combined with thermophilic fungus compared to treatments contained non-heated microsclerotia 75 days after sowing (Table 3). A increase in leaf number was also observed in treatments containing microsclerotia heated under program 3 combined with thermophilic fungus and VAM spores compared to treatments contained microsclerotia heated under program 3 (without thermophillic fungus). Inoculation of soil with VAM spores caused an additional decrease in disease incidence 30 days after planting seeds (Table 4). Two months after sowing, a significant increase in leaf infection index was observed only in treatment contained microsclerotia heated under program 2 and thermophilic fungus spores (Table 4). Seventy five days after planting seeds, there was the significant increase of the leaf infection index mean in treatments contained non-heated microsclerotia and microsclerotia heated under program 1 separately compared to all treatments except treatments contained microsclerotia heated under other two programs (Table 4). The stability process of the significant increase of the leaf infection index showed that during three months after sowing, treatments contained microsclerotia heated under different programs separately combined with both thermophilic fungus and VAM spores were not belonged to treatment groups with the significant increase of the leaf infection index (Table 4). Appearance of the significant different of the leaf infection index in treatments contained non-heated and heated microsclerotia combined with thermophilic fungus and VAM spores was observed 30 days after planting seeds and continued until 2 months after planting seeds however there was the significant increase of the leaf infection index in treatments contained non-heated and heated microsclerotia 30 days after planting seeds and this manner was continued until 75 days after planting seeds (Table 4). In studies concerned to plant weight, there was the significant increase of the plant weight mean in treatment contained microsclerotia heated under program 3 combined with both thermophilic fungus and VAM spores compared to other treatments (Table 5). Thus, the significant increase of the plant weight was observed in all treatments contained both thermophilic fungus and VAM spores compared to all treatments contained single VAM spores.
| Table 1: | The stability process of the significant increase of the leaf surface mean in some treatments compared to other treatments during 3 months after seeds planting1 |
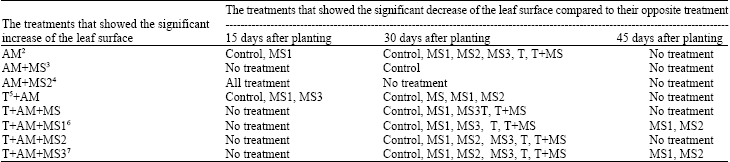 | |
1There was not any significant different between the leaf surface means of the different treatments 60, 75 and 90 days after planting. 2AM: treatment contained VAM spores; 3MS: treatment contained non-heated microsclerotia; 4MS2: treatment contained microsclerotia heated under program2; 5T: treatment contained thermophilic fungus spores; 6MS1: treatment contained microsclerotia heated under program1; 7MS3: treatment contained microsclerotia heated under program3 | |
| Table 2: | The stability process of the significant increase of the plant height mean in some treatments compared to other treatments during 3 months after seeds planting1 |
 | |
There was not any significant different between the plant height means of the different treatments 60, 75 and 90 days after planting | |
| Table 3: | The stability process of the significant increase of the leaf number mean in some treatments compared to other treatments during 3 months after seeds planting1 |
 | |
1There was not any significant different between the leaf number means of the different treatments 15, 30 , 45 and 90 days after planting | |
| Table 4: | The stability process of the significant increase of the leaf infection index mean in some treatments compared to other treatments during 3 months after seeds planting1 |
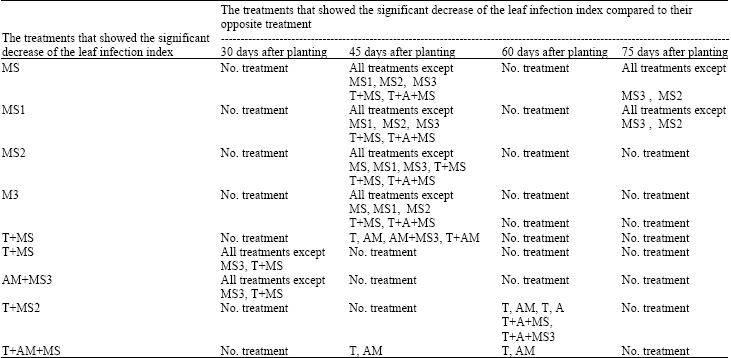 | |
1There was not any significant different between the disease severity index means of the different treatments 15 and 90 days after planting | |
| Table 5: | Statistical grouping different treatments in order to determining the significant different of dry weight mean in the end of a vegetative period |
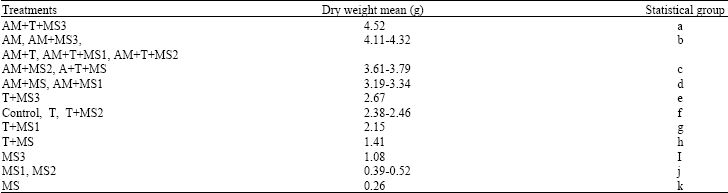 | |
| Table 6: | The appointment of the correlation between the several of the vegetative qualities (plant height, leaf number and leaf surface) and leaf infection index during 3 months after seeds planting in different treatments |
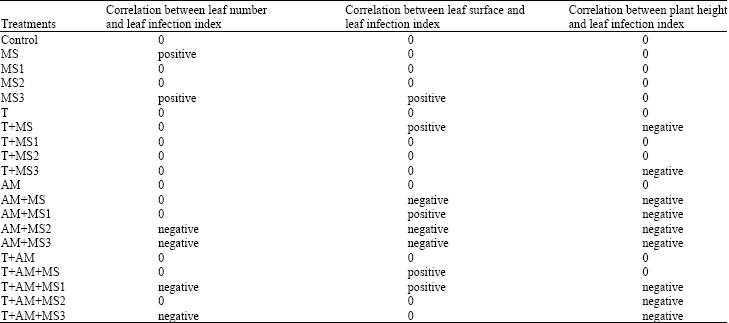 | |
On the other hand, there was the significant increase of the plant weight mean in all treatments contained single VAM spores compared to all treatments contained single thermophilic fungus spores. Thus, the significant increase of the plant weight was observed in all treatments contained single thermophilic fungus spores compared to all treatments without spores. There was the significant increase of the plant weight in all treatments contained microsclerotia heated under program 3 compared to similar treatments but contained microsclerotia heated under two other programs. The microsclerotia heated under program 3 resulted in higher plant dry weight compared to those of heated under program 1 in treatments contained thermophilic fungus and VAM spores individually. Thus, the microsclerotia heated under program 1 resulted in higher plant dry weight in treatments contained single thermophilic fungus spores and combined with VAM spores compared to those of non-heated. The results of this research showed that there was a direct relation between the number of leaves and leaf infection index in treatments contained microsclerotia heated under different programs and non-heated. There was a reversed relation between the number of leaves and leaf infection index in some treatments contained microsclerotia heated under different programs combined with either single VAM spores or both VAM and thermophilic fungus spores. The results of the studies concerned to correlation between leaf surface and leaf infection index showed that there was a reversed relation in all treatments contained single VAM spores. Thus, there was a reversed relation between plant height and leaf infection index in all treatments contained VAM and thermophilic spores individually or together (Table 6).
DISCUSSION
The overall results of this study show that it may be possible to manage Verticillium wilt disease of cotton efficiently by using a combination of strategies such as soil heat treatment, thermophillic fungus and VAM compared to use of each of these factors separately. The results of our study were similar to those of Tjamos and Fravel (1995) in some extend but there were also some differences in the impact of treatments on plant growth characteristics such as leaf surface area and plant height.
The symptoms of Verticillium wilt diseases usually appear after a relatively long period of time. In this study disease symptoms appeared on leaves 45 days after sowing. This is perhaps due to the primary resistance and responses of the plant to pathogen attack and infection. Plants use several defense mechanisms against pathogenic agents attack including production of phytoalexins (Yoshikawa et al., 1993). These low molecular weight compounds play important roles in plant natural defense mechanism (Nawar and Kuti, 2003). On the other hand, plant pathogens also use different mechanism to attack and infect host plants (Morrissey and Osbourn (1999). Production and secretion of different enzymes such as pectinase and endopolygalactronase (Misaghi, 1982) is one of theses mechanisms. The interactions among the above mentioned plant and pathogen strategies result in susceptibility or resistance of plants to diseases.
In this study, the increase in the plant height, number of leaves and leaf surface area and the decrease of leaf infection index were significant in all treatments contained both VAM and thermophilic fungus spores compared to the treatments without these spores 4 to 8 weeks after sowing. In a previous study by Tjamos and Fravel (1995). It was found that there were significant decrease in leaf infection index in all treatments contained microsclerotia heated compared to all treatments contained microsclerotia non- heated (Tjamos and Fravel, 1995). A common subject concerned to function of plants, is that plant products in generative period depend on plant biomass in vegetative period (Chailakhyan, 1968). It is conceived that the increase of the plant height is performed a reveal impression in disappearing diseases symptoms.
In this study, the dry weight of plants in treatments contained VAM was higher than treatments contained thermophilic fungus spores and minimum dry weight was found in treatments without VAM and thermophilic fungus. Present results also indicate that there was significant differences in dry weight of plants among treatments contained microsclerotia heated under different programs. Maximum dry weight of plant was concerned to treatments contained heated microsclerotia under program 3 (35°C for 10 h followed by 38°C for 14 h). The increase in the absorption of nutrients induced by VAM w could increase plant resistance against pathogenic agents and this has been shown in previous studies (Linderman, 1994).
Overall results of this show that the use of thermophilic fungus, VAM and heat treatment together could be effective strategies in controlling Verticillium wilt which is an important disease of cotton and several other crops (Naraghi, 1999). Present findings may have practical application in disease management in the field. Present study was conducted in the greenhouse condition to achieve fast primary results in approving our hypothesis. For further and complete investigation of this subject more experiments may be conducted in field condition to come up with an applied instruction for management of Verticillium wilt disease on cotton and other crops.
REFERENCES
- Atkinson, D., G. Berta and J.E. Hooker, 1994. Impact of Mycorrhizal Colonization Root Architecture, Root Longevity and the Formation of Growth Regulators. In: Impact of Arbuscular Mycorrhizas on Sustainable Agriculture Natural Ecosystems, Gianinazzi, S. and H. Schuepp (Eds.). Birkhauser Verlag, Basel, pp: 89-99.
- Aziz, N.H., M.Z. El-Fouly, A.A. El-Essawy and M.A. Khalaf, 1997. Influence of bean seedling root exudates on the rhizosphere colonization by Trichoderma lignorum for the control of Rhizoctonia solani. Bot. Bull. Acad. Sinica, 38: 33-39.
Direct Link - Borowicz, V.A., 2001. Do arbuscular mycorrhizal fungi alter plant-pathogen relations. Ecology, 82: 3057-3068.
Direct Link - Fortuna, P., A.S. Citernesi, S. Morini, C. Vitagliano and M. Giovannetti, 1996. Influence of arbuscular mycorrhiza and phosphate fertilization on shoot apical growth of micropropagated apple and plum rootstocks. Tree Physiol., 16: 757-763.
Direct Link - Fortuna, P., S. Morini and M. Giovannetti, 1998. Effects of arbuscular mycorrhizal fungi on in vivo root initiation and development of micropropagated plum shoots. J. Hortic. Sci. Biotechnol., 73: 19-28.
Direct Link - Gavito, M.E. and P.A. Olsson, 2003. Allocation of plant carbon to foraging and storage in arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol., 45: 181-187.
Direct Link - Heydari, A., I.J. Misaghi and W.B. McCloskey, 1997. Effects of three soil-applied herbicides on populations of plant disease suppressing bacteria in the cotton rhizosphere. Plant Soil, 195: 75-81.
CrossRefDirect Link - Heydari, A. and I.J. Misaghi, 1998. The impact of herbicides on the incidence and development of Rhizoctonia solani-induced cotton seedling damping-off. Plant Dis., 82: 110-113.
Direct Link - Heydari, A. and I.J. Misaghi, 2003. The role of rhizosphere bacteria in herbicide-mediated increase in Rhizoctonia solani-induced cotton seedling damping-off. Plant Soil, 257: 391-396.
CrossRefDirect Link - Hwang, S.F., K.F. Chang and P. Chakravarty, 1992. Effects of vesicular-arbuscular mycorrhizal fungi on the development of Verticillium and Fusarium wilts of alfalfa. Plant Dis., 76: 239-243.
CrossRef - Linderman, R.G. and A.A. Davis, 2001. Comparative response of selected grapevine rootstocks and cultivars to inoculation with different mycorrhizal fungi. Am. J. Enol. Viticult., 52: 8-11.
Direct Link - Marois, J.J., D.R. Fravel and G.C. Papavizas, 1984. Ability of Talaromyces flavus to occupy the rhizosphere. Soil Biol. Biochem., 16: 387-390.
Direct Link - Morrissey, J.P. and A.E. Osbourn, 1999. Fungal resistance to plant antibiotics as a mechanism of pathogenesis. Microbiol. Mol. Biol. Rev., 63: 708-724.
Direct Link - Murray, F.R., D.J. Llewellyn, W.J. Peacock and E.S. Dennis, 1997. Isolation of the glucose oxidase gene from Talaromyces flavus and characterization of its role in the biocontrol of Verticillium dahliae. Curr. Genet., 35: 367-375.
Direct Link - Naraghi, L., F. Azad-Disfani and A. Heydari, 2004. The possibility of Talaromyces flavus using in biological control on Verticillium wilt of cotton. Proceedings of the 3rd National Conference on the Development in the Application of Biological Products and Optimum Utilization of Chemical Fertilizer and Pesticides in Agriculture, February 21-23, 2004, Tehran, Iran, pp: 411.
- Nawar, H.F. and J.O. Kuti, 2003. Wyerone acid phytoalexin synthesis and peroxidase activity as markers for resistance of broad beans to chocolate spot disease. J. Phytopathol., 151: 564-570.
CrossRefDirect Link - Liu, R.J., 1995. Effect of vesicular-arbuscular mycorrhizal fungi on Verticillium wilt of cotton. Mycorrhiza, 5: 293-297.
Direct Link - Spink, D. and R.C. Rowe, 1989. Evaluation of Talaromyces flavus as a biological control agent against Verticillium dahliae in potato. Plant Dis., 73: 230-236.
CrossRefDirect Link - Tjamos, E.C. and D.R. Fravel, 1995. Detrimental effects of sublethal heating and Talaromyces flavus on microsclerotia of Verticillium dahliae. Phytopathology, 85: 388-392.
Direct Link - Yoshikawa, M., M. Tsuda and Y. Takeuchi, 1993. Resistance to fungal diseases in transgenic tobacco plants expressing the phytoalexin elicitor-releasing factor, B-1, 3-endoglocanase, from Soybean. Naturwissenschaften, 80: 417-420.
CrossRefDirect Link