
Tawatchai Tanee
Department of Biochemistry, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Arunrat Chaveerach
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Nison Sattayasai
Department of Biochemistry, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Alongkoad Tanomtong
Department of Biology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Scott A. Suarez
Department of Anthropology, Miami University, USA
Suporn Nuchadomrong
Department of Biochemistry, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 9 | Page No.: 1459-1464







ABSTRACT
Cytogenetic studies of three gibbon species using conventional banding patterns were investigated showing an identical number of 44 diploid chromosomes. They are assumed to have common evolutionary relationships. For in depth study, molecular markers were assessed using the Amplified Fragment Length Polymorphism (AFLP) method. With seven successful primer combinations, a total of 1669 scorable bands were generated. The resulting bands were used for dendrogram construction. From the dendrogram, the individuals of Hylobates lar are closely related to H. agilis more than H. pileatus with a bootstrap value of 78%. Averages of inter-specific genetic similarity values among all gibbon species studied are 67.15% (between H. lar and H. pileatus) to 71.03% (between H. lar and H. agilis). In summary three gibbon species show genetic stability within a species. The development of specific molecular markers of a species is beneficial for genetic differentiation of this group of primates.
PDF Abstract XML References Citation
How to cite this article
Tawatchai Tanee, Arunrat Chaveerach, Nison Sattayasai, Alongkoad Tanomtong, Scott A. Suarez and Suporn Nuchadomrong, 2007. Analysis of the Genetic Relationships among Thai Gibbon Species Using AFLP Markers. Pakistan Journal of Biological Sciences, 10: 1459-1464.
DOI: 10.3923/pjbs.2007.1459.1464
URL: https://scialert.net/abstract/?doi=pjbs.2007.1459.1464
DOI: 10.3923/pjbs.2007.1459.1464
URL: https://scialert.net/abstract/?doi=pjbs.2007.1459.1464
INTRODUCTION
The hylobatidae (gibbons) are a relatively small and morphologically homogeneous group of primate species living in closed canopy rain forest throughout Southeast Asia (Takacs et al., 2005). They consist of 12 species and are subdivided into 4 morphologically and karyologically distinct genera, namely Hylobates (diploid number (2n) = 44) containing H. lar, H. pileatus, H. agilis, H. moloch, H. muelleri and H. klossii, Hoolock (2n = 38) containing H. hoolock, Nomascus (2n = 52) containing N. concolor, Nomascus sp. cf. nasutus, N. gabriellae and N. leucogenys and Symphalangus (2n = 50) containing S. syndactylus (Geissmann, 2002). Three of these have been seen in Thailand including H. lar, H. pileatus and H. agilis that are only in the lar group.
Scientists have little understanding of the biogeographic history of gibbons, largely because of their sparse fossil record (Geissman, 2002; Takacs et al., 2005). A limited understanding of the environmental history of Thailand further compounds the problem. Researchers have proposed few scenarios to describe the radiation of gibbons. Moreover, habitat loss and fragmentation, habitat degradation, hunting (food, medicine and sport) and illegal trade (medicine, pets) are the top four which have seriously threatened genetic resources of gibbons throughout their ranges.
The genetic relationships among different gibbon species are still controversially discussed with limited material. The currently accepted classification of gibbons is mainly based on morphological studies (Geissmann, 2002), while their cytogenetics and molecular genetics have become important tools (Tanomtong et al., 2005; 2006; Chaveerach et al., 2006; Tanee et al., 2006). As part of a study on the complete genetic relationships of Hylobatidae, Thai gibbons play a key role, because they are endangered species and the small amount of available data.
The Amplified Fragment Length Polymorphism (AFLP) technique is one DNA fingerprinting procedure which uses PCR to amplify a limited set of DNA fragments from a specific DNA sample. Since AFLP analysis does not require prior genetic information of the taxa studied, it should be of value in phylogenetic analysis of a wide variety of organisms (Vos et al., 1995). Furthermore, AFLP phenotypes are highly reproducible (Ovilo et al., 2000; Liu et al., 2005) and thus, reliable (Aggarwal et al., 1999; Sudupak et al., 2004). The AFLP markers have been widely applied in organisms for analyzing genetic variation and phylogenetic relationships among and within closely related species (Aggarwal et al., 1999; Mace et al., 1999; Loh et al., 2000; Despres et al., 2002; Pelser et al., 2003; Ude et al., 2003; Wang et al., 2004; Banfer et al., 2004; Chen et al., 2004; Sudupak et al., 2004); Liu et al., 2005).
In this study we investigated the genetic relationships among H. agilis, H. lar and H. pileatus that are in the lar group using the AFLP technique in order to clarify the relationships among these three gibbon species.
MATERIALS AND METHODS
Sample collection: Blood samples from 2 black handed gibbons (H. agilis from Southern Thailand), 5 white handed gibbons (H. lar Northeastern Thailand) and 3 pileatus gibbons (H. pileatus from Northeastern Thailand) were collected (Fig. 1). The samples were divided into two groups, the first for cytogenetic analysis and the second for molecular analysis.
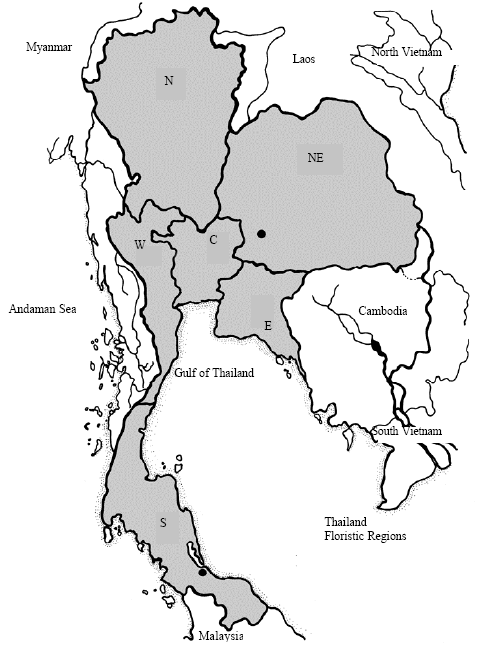 | |
| Fig. 1: | Map of Thailand indicating sample collection of gibbons (•), where N is Northern Thailand, NE is Northeastern Thailand, C is Central Thailand, W is Western Thailand, E is Eastern Thailand and S is Southern Thailand |
Cytogenetic analysis: Blood samples were subjected to cytogenetic studies by lymphocyte culture of whole blood. The cultured cells were examined by the colchicines-hypotonic fixation-air drying technique followed by a conventional technique (Tanomtong et al., 2005). Chromosomal checks were performed with 20 cells of each individual by light microscopy.
Molecular analysis
DNA isolation: Genomic DNA was extracted from blood samples following Sambrook and Russel (2001). The quality and quantity of extracted DNA was assessed by 0.8% agarose gel electrophoresis and spectrophotometry.
AFLP reaction: The procedures of AFLP method (Vos et al., 1995) were performed according to the protocol of the Kit (AFLPR Analysis System I, Invitrogen, USA). After adaptor ligation and preselective amplification, selective amplification was conducted with seven primer combinations: E-ACC/M-CAG, E-AAC/M-CAT, E-AAC/M-CAG, E-AAG/M-CAT, E-AAG/M-CTT, E-AGG/M-CTT and E-AGC/M-CAT. The PCR products amplified with different primer combinations were loaded onto 6.0% denaturing polyacrylamide gels and electrophoresed for 3 h and detected by Silver QuestTM Silver Staining Kit (Invitrogen, USA).
Data analysis: Each AFLP band was considered as an independent character and the bands were scored visually as having each band either absent (0) and present (1) across all samples with the same primer pairs. Qualitative differences in band intensity were not considered. With the band data, a pair-wise genetic similarity matrix was generated among Hylobates lar, H. agilis and H. pileatus individuals using Ochiai similarity coefficients which were then converted to a genetic distance matrix. Based on the genetic distance matrix, cluster analyses were performed and the corresponding dendrograms were constructed for the three gibbon species using the Single Linkage Cluster method. Cophenetic correlations were computed from the clustering matrix in order to get the best fit dendrogram. All of these analyses were performed using the Fingerprinting II program (BioRad, USA).
RESULTS
Results from lymphocyte culture of whole blood and conventional staining of the three gibbons species, namely Hylobates lar, H. agilis and H. pileatus indicate that all species have an identical number of 44 diploid chromosomes (2n) consisting of 42 autosomes and 2 sex chromosomes. A representative chromosome of the three gibbon species is from a male of H. pileatus as shown in Fig. 2.
AFLP fragments were established for five individuals of H. lar, two individuals of H. agilis and three individuals of H. pileatus. Seven primer combinations generated a total of 1669 scorable bands, averaging at 238 bands per primer. Of these bands, 32.59% (544 bands) were polymorphic. Figure 3 presents AFLP fingerprinting of three gibbons with seven primer combinations.
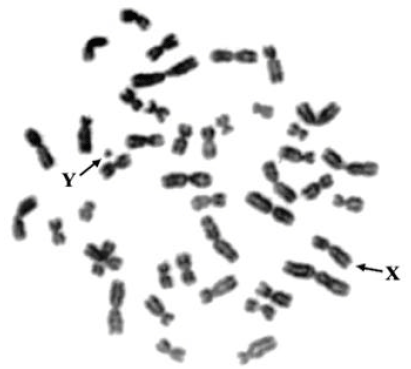 | |
| Fig. 2: | Diploid chromosome number of male H. pileatus, 2n = 44 consisting of 42 autosomes and 2 sex chromosomes |
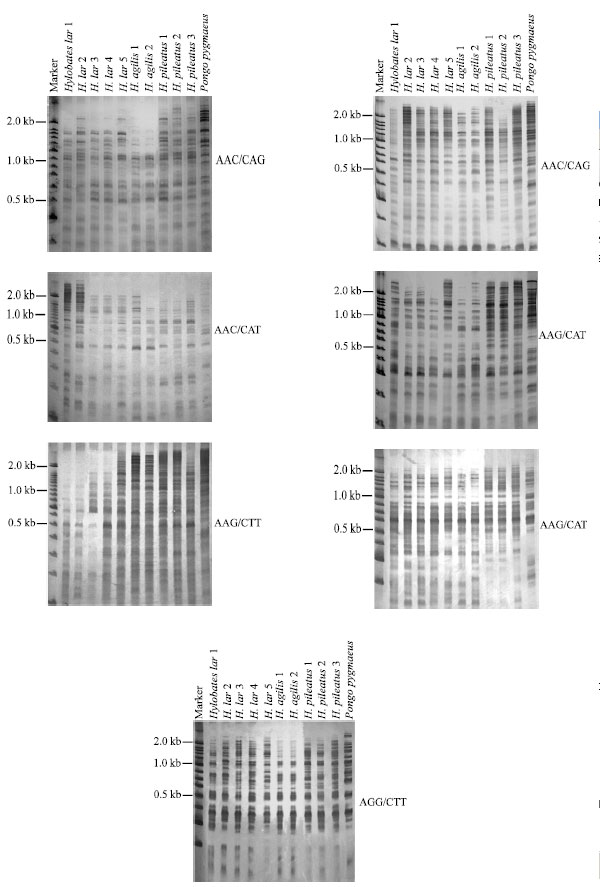 | |
| Fig. 3: | Specific banding pattern of three gibbon species with the seven successful primer combinations |
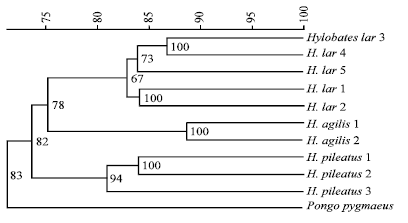 | |
| Fig. 4: | Dendrogram depicting the seven AFLP primer combinations produced by single linkage cluster analysis that are used to clarify the genetic relationships among the three gibbon species |
| Table 1: | Average similarity index values based on AFLP analysis of Hylobatid species and P. pygmaeus |
 | |
A dendrogram was constructed based on the AFLP scoring data and using single linkage cluster analysis. The three hylobatid species are clustered together, whereas Pongo pygmaeus is separated as an outgroup with a bootstrap value of 83%. Within the hylobatid cluster, H. lar is more closely related to H. agilis than H. pileatus (Fig. 4). Average values of inter-specific similarity index (S) among all hylobatid studied species are 67.15% (between H. lar and H. pileatus) to 71.03% (between H. lar and H. agilis) (Table 1).
DISCUSSION
The identity of diploid chromosome number (2n = 44) of three gibbon species agrees with Ankel-Simons (2000) and Geissmann (2002), reflecting the close genetic relationships among species of gibbons. Moreover, Ankel-Simons (2000) suggested that chromosome polymorphism and inter-specific chromosomal variability are common among prosimian primates. Primate cytogenetics can still not reconstruct reliably the relatedness of gibbon species. The AFLP molecular marker was thus used to elucidate their relatedness. The advantage of AFLP fingerprints is that they are highly reproducible in the samples studied.
The sample studied contained a small number of some species such as H. agilis because blood samples are difficult to collect from primates in the forest. However, at least two samples could be used for preparing DNA fingerprints. Within the Hylobates species (Fig. 4), H. lar is very closely related to H. agilis more than H. pileatus with high bootstrap value (78%) and high average S-value (71.03%). This result is supported by other work using DNA sequences (Garza and Woodruff, 1992; Hayashi et al., 1995; Takacs et al., 2005), morphology (Kimble and Martin, 1993) and by vocal data (Geissmann, 2002).
Based on the geography of the Khao Yai National Park, Thailand, two members of the lar group, H. lar and H. pileatus, meet and occasionally hybridize. In the Khao Yai overlap zone, pure H. lar can be found 5 km into the pileatus side of the zone and pure H. pileatus go 9-12 km into the lar side. The change of the morphological index from 90% lar to 90% pileatus occurs over about 9 km (Kimbel and Matin, 1993). However, the genetic relationships between H. lar and H. pileatus is low with a 67.15% S value, while the S value between H. agilis and H. pileatus is 69.97%. Therefore, we suggest that although H. lar and H. pileatus have overlapping habitation zones and habitat has been lost, they still show genetic stability within a species. In order to resolve the genetic relationships within the lar group properly, it is necessary to study a larger sample and cover distributed areas, as well as to consider comparative behavioral studies. Thus, these results suggest that genetic analysis based on AFLP fingerprinting has a good capacity for study of species diversity, especially in gibbons. As AFLP analysis does not require prior genetic information of the taxa studied and AFLP fingerprints are reproducible, the technique should be of value in genetic analysis of a wide variety of primates (Chaveerach et al., 2006; Tanee et al., 2006). Moreover, the seven successful primer combinations can be used to sufficiently determine genetic differences among gibbon species.
ACKNOWLEDGMENT
This work was supported in part by Khon Kaen University’s Graduate Research Fund, Academic Year 2004.
REFERENCES
- Banfer, G., B. Fiala and K. Weising, 2004. AFLP analysis of phylogenetic relationships among myrmecophytic species of Macaranga (Euphorbiaceae) and their allies. Plant Sys. Evol., 249: 213-231.
CrossRefDirect Link - Chaveerach, A., T. Tanee, N. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2007. Genetic relationships of langur species using AFLP markers. Pak. J. Biol. Sci., 10: 1445-1451.
CrossRefPubMedDirect Link - Chen, J., P.S. Devanand, D.J. Norman, R.J. Henny and C.C.T. Chao, 2004. Genetic relationships of aglaonema species and cultivars inferred from AFLP Markers. Ann. Bot., 93: 157-166.
Direct Link - Despres, L., L. Gielly, B. Redoutet and P. Taberlet, 2003. Using AFLP to resolve phylogenetic relationships in a morphologically diversified plant species complex when nuclear and chloroplast sequences fail to reveal variability. Mol. Phylogen. Evol., 27: 185-196.
CrossRefDirect Link - Liu, Y.G., S.L. Chen, B.F. Li, Z.J. Wang and Z. Liu, 2005. Analysis of genetic variation in selected stocks of hatchery flounder, Paralichthys olivaceus, using AFLP markers. Biochem. Syst. Ecol., 33: 993-1005.
Direct Link - Loh, J.P., R. Kiew, O. Set, L.H. Gan and Y.Y. Gan, 2000. Amplified fragment length polymorphism fingerprinting of 16 banana cultivars (Musa cvs.). Mol. Phylogene. Evol., 17: 360-366.
CrossRefDirect Link - Mace, E.S., C.G. Gebhardt and R.N. Lester, 1999. AFLP analysis of genetic relationships in the tribe Datureae (Solanaceae). Theor. Applied Genet., 99: 634-641.
CrossRefPubMedDirect Link - Ovilo, C., M.T. Cervera, C. Castellanos and J.M. Martinez-Zapater, 2000. Characteriztion of Iberian pig genotypes using AFLP markers. Anim. Gene., 31: 117-122.
PubMedDirect Link - Pelser, P.B., B. Gravendeel and R.V.D. Meijden, 2003. Phylogeny reconstruction in the gap between too little and too much divergence: The closest relatives of Senecio jacobaea (Asteraceae) according to DNA sequences and AFLPs. Mol. Phylogene. Evol., 29: 613-628.
Direct Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link - Sudupak, M.A., M.S. Akkaya and A. Kence, 2004. Genetic relationships among perennial and annual Cicer species growing in Turkey assessed by AFLP fingerprinting. Theor. Applied Gene., 108: 937-944.
Direct Link - Takacs, Z., J.C. Morales, T. Geissmann and D.J. Melnick, 2005. A complete species-level phylogeny of the hylobatidae based on mitochondrial ND3-ND4 gene sequences. Mol. Phylogene. Evol., 36: 456-467.
Direct Link - Tanee, T., A. Chaveerach, A. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2006. Cytogenetic and AFLP fingerprints for five species of Thai Macaques. Cytologia, 713: 303-308.
CrossRefDirect Link - Ude, G.N., W.J. Kenworthy, J.M. Costa, P.B. Cregan and J. Alvernaz, 2003. Genetic diversity of soybean cultivars from China, Japan, North America and North American ancestral lines determined by amplified fragment length polymorphism. Crop Sci., 43: 1858-1867.
CrossRefDirect Link - Vos, P., R. Hogers, M. Bleeker, M. Reijans and T. van de Lee et al., 1995. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res., 23: 4407-4414.
CrossRefPubMedDirect Link