
M. D. Allarangaye
lnstitut Tchadien de Recherche Agronomique Pour le Developpement (ITRAD),
BP 5400 N`jamena, Tchad
O. Traore
lnstitut de l`Environnement et de Recherches Agricoles (INERA),
01 BP 476 Ouagadougou 01, Burkina Faso
E.V. S. Traore
UFR/SVT, Departement de Biologie et Ecologie V egetales, Universite de Ouagadougou,
03 BP 7021 Ouagadougou 03, Burkina Faso
R. J. Millogo
lnstitut de l`Environnement et de Recherches Agricoles (INERA),
01 BP 476 Ouagadougou 01, Burkina Faso
S. Guinko
UFR/SVT, Departement de Biologie et Ecologie V egetales, Universite de Ouagadougou,
03 BP 7021 Ouagadougou 03, Burkina Faso
G. Konate
UFR/SVT, Departement de Biologie et Ecologie V egetales, Universite de Ouagadougou,
03 BP 7021 Ouagadougou 03, Burkina Faso
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 9 | Page No.: 1414-1421







ABSTRACT
In the present study, we investigated on the experimental host range of RYMV among plant species most of which are frequently encountered in rice-growing environments of west and central African savannahs. Only seven out of 66 plant species inoculated were infected by RYMV. All susceptible plant species belonged to the Poaceae family and three of them (Chloris prieuri, Eragrostis cilianensis and Shoenefeldia gracilis) were reported for the first time. Symptoms were conspicuous and persistent in most species but disappeared totally in older plants of some host species such as S. gracilis and Eragrostis tenella. Therefore, surveys for identification of RYMV wild hosts should be conducted before the flowering stage. Virus-host Interactions were studied between 15 RYMV isolates of different strains and 10 wild host species. Differential reactions were obtained in the crow-foot grass Dactyloctenium aegyptium which was susceptible to five of the fifteen isolates. All other plants were susceptible to the whole set of virus isolates. Altogether, this study underlined the narrowness of RYMV host range and pointed out the complexity of interactions between the virus and its hosts, especially the rationale behind overcoming host barriers.
PDF Abstract XML References Citation
How to cite this article
M. D. Allarangaye, O. Traore, E.V. S. Traore, R. J. Millogo, S. Guinko and G. Konate, 2007. Host Range of Rice yellow mottle virus in Sudano-Sahelian Savannahs. Pakistan Journal of Biological Sciences, 10: 1414-1421.
DOI: 10.3923/pjbs.2007.1414.1421
URL: https://scialert.net/abstract/?doi=pjbs.2007.1414.1421
DOI: 10.3923/pjbs.2007.1414.1421
URL: https://scialert.net/abstract/?doi=pjbs.2007.1414.1421
INTRODUCTION
Rice (Oryza sativa L.) is a major staple crop in Africa where it is used for feeding both human and cattle. Improving its production is one of the main goals for reaching more food security and alleviating recurrent food shortages in different parts of the continent. However, since the early 1990s, rice production is severely affected by rice yellow mottle, the most important viral disease of rice in Africa. The disease occurs in most countries south of the Sahara and also in the islands of Zanzibar and Madagascar (Kouassi et al., 2005). The causal agent is Rice yellow mottle virus (RYMV) of the plant virus genus Sobemovirus (Hull and Fargette, 2005). Depending on the rice cultivar, RYMV induces various symptoms, including yellow or orange discoloration of the leaves, reduced tillering, stunting of plants and sterility of flowers. During epidemics, yield can be lost completely, especially when susceptible rice cultivars are grown and plants infected at an early stage of development (Calvert et al., 2003).
As a major contribution to the genetic control of rice yellow mottle, resistance to RYMV has been found in rice cultivar Gigante (O. sativa indica) and in some cultivars of the African rice O. glaberrima Steud. such as Tog 5681, Tog 5672 and Tog 5675. However, the success of using this resistance is undermined by the occurrence of highly pathogenic strains of RYMV that are capable to overcome the resistance (Traoré et al., 2006a).
Large scale studies of RYMV isolates also indicated the occurrence of several serotypes and phylogenetic strains with a geography-based distribution (Pinel et al., 2000; Fargette et al., 2004; Traoré et al., 2005). Three serotypes named Ser1, Ser2 and Ser-Sa were identified in West and Central African savannahs (Traoré, 2006). Two other serotypes including Ser4 and Ser5 were found in East Africa and in Madagascar (Fargette et al., 2002). In general, RYMV serotypes could be assigned to corresponding phylogenetic strains based on coat protein gene sequence analysis. However, Ser 1 isolates of West Africa and those of Central Africa were found to belong to two phylogenetic sister groups named S1-WA and S1-CA, respectively (Traoré et al., 2005).
RYMV is a non seed-transmitted virus (Fauquet and Thouvenel, 1977; Konaté et al., 2001; Allarangaye et al., 2006) but it is readily transmitted by mechanical inoculation (Bakker, 1970). RYMV is also transmitted through several other means, including insect vectors (mainly beetles) (Bakker, 1974; Heinrichs et al., 1997; Banwo and Makundi, 2001), mammals like cows, donkeys and rats (Sarra and Peters, 2003), wind through contact between plants (Sarra et al., 2004) and cropping practices involving the use of seedbeds (Traoré et al., 2006b). The role of some of these factors in rice yellow mottle epidemiology is straightforward. Seedbeds are likely involved only in the primary spread of the virus in the field. Alternatively, the action of wind is to be placed within the secondary spread of the virus from already established infections. Factors such as mammals or insect vectors may play a role in both primary and secondary spread of the disease.
Sources of inoculum for primary infections to occur in seedbeds or in rice fields are not fully known. Infected rice stubble from previously harvested crops and wild host plants acting as reservoirs for RYMV, have been reported as major inoculum sources (Abo et al., 2000). The natural host range of RYMV is still poorly known and limited to wild rice species O. longistaminata Chev. and Roehr and O. barthii A. Chev and to a few wild reservoir hosts including Ischaemum rugosum Salisb, Echinochloa colona Link, Eragrostis atrovirens (Desf.) Trin. ex Steud and Panicum repens L. (Konaté et al., 1997; Abo et al., 2000; Traoré, 2006). Consequently, the role of wild hosts in rice yellow mottle epidemiology has not been clearly determined. Identification of RYMV host plant species is made difficult because some of them i.e., Eragrostis tenella may sustain asymptomatic though systemic infections (Bakker, 1974). Therefore, it is advisable to first determine the experimental host range of the virus in order to make the identification of natural hosts easier. An extensive study of the host range of RYMV was conducted in East Africa by inoculation of the virus to test plants (Bakker, 1974). Apart from rice and its related wild species, only seven systemic hosts were identified, most of which belonged to the Eragrostideae tribe of the plant family Poaceae.
There have been some conflicting results between RYMV host range studies conducted in East and West Africa. Plant species such as Echinochloa colona, Eragrostis atrovirens and Panicum repens reported to be hosts of RYMV in West Africa (Awoderu, 1991; Konaté et al., 1997; Traoré, 2006) were found to be non-host species in East Africa (Bakker, 1974). Moreover, preliminary results on reactions of some host plants with virus isolates from West and Central Africa did not fully agree with Bakker’s studies in East Africa. For example, the crows-foot grass Dactyloctenium aegyptium (L.) Beauv. was found to be a systemic host that exhibited clear mottle symptoms when challenged with a Chadian isolate of RYMV (Allarangaye et al., 2006). In the contrary, when this species was challenged with the East African isolate, only local lesions in which virus particles could be recovered irregularly were induced (Bakker, 1974).
In the present study, we investigated on the experimental host range of RYMV among plant species most of which are frequently encountered in rice-growing environments of west and central African savannahs. Implications for the identification of natural hosts and their role in rice yellow mottle epidemiology are discussed.
MATERIALS AND METHODS
Sudano-Sahelian zone and virus isolates: The Sudano-Sahelian zone located in Sub-Saharan Africa is a strip which stretches from Senegal to Sudan and is enclosed by 7° and 15° North latitude and 17° West and 24° East longitude. This area is semi-arid and includes three major agroclimatic zones in a north to south direction: (a) Sahelian zone bordered by isohyets 600 mm to the south and 300 mm the north; (b) Sudan savannah zone located between isohyets 600 mm to the north and 900 mm to the south; (c) Northern guinea savannah with 900-1200 mm of rainfall par year. The virus isolates used in this study were from our virus collection and originated from four countries of West (Burkina Faso and Mali) and Central Africa (Chad and Cameroon). RYMV diversity in these areas was taken into account and the isolates were chosen in relation to the serotype and pathotype they belonged to. Serotypes were differentiated by using anti-RYMV monoclonal antibodies mab A and mab D (Traoré, 2006). Isolates of serotype Ser1 reacted with mab A only. S2 were solely detected by mab D whereas Ser-Sa reacted with both antibodies. RYMV pathotypes were defined according the ability of isolates to induce symptoms or not in resistant rice cultivars Gigante and Tog 5681 (Table 1).
Host range studies: Virus isolates were first propagated in 2 week-old plants of the susceptible rice cultivar BG90-2 by mechanical inoculation. Symptoms developed fully at 14 days post-inoculation (dpi) and infected leaves were collected. Portions of 0.1 g of leaves containing each virus isolate were taken and ground together with sterile mortars and pestles in 10 mL of 100 mM phosphate buffer, pH 7.0. Carborundum (600 mesh) was added to the homogenate which was subsequently rubbed onto leaves of test plants which included Poaceae species mainly and also several dicotyledonous and other monocotyledonous species.
| Table 1: | Origin and properties of RYMV isolates used in this study |
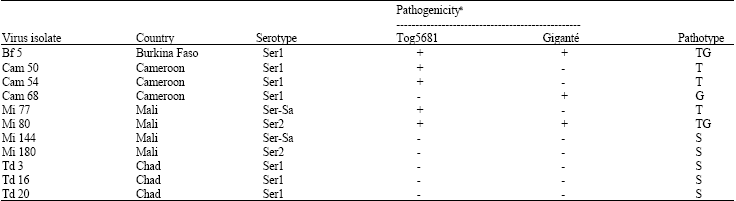 | |
| aCapability of RYMV isolates to induce symptoms (+) or not (-) in the resistant rice cultivars Tog5681 and Gigante | |
A few plant species (mainly of the genus Eragrostis) which were subject to contrasting identification as hosts of RYMV (Allarangaye et al., 2006; Bakker, 1974) were also tested. Most plant species tested are widespread in rice-growing environments of West and Central African savannahs (Johnson, 1997; Akobundu and Agyakwa, 1989). Test plants were grown in 1-litre-plastic pots from seeds for most species but cuttings were used for a few species which reproduce vegetatively. Plants were inoculated at 3-5 leaf stage and maintained in an insect-proof greenhouse at 25-30°C and relative humidity of 80-90%. Appearance of symptoms was monitored during 45 dpi and leaf samples were collected for virus detection and back inoculation tests.
Serological and back inoculation tests: Double antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) was used to detect RYMV in leaf samples (Clark and Adams, 1977). A broad spectrum polyclonal antibody to major RYMV strains of West and Central Africa was used as the coating antibody (Traoré, 2006). The same antibody was coupled to alkaline phosphatase and used as conjugate. All buffer systems and incubation times were as previously described (Konaté et al., 1997). Extracts used as sources of antigens were obtained by grinding the leaf samples (1 g in 10 mL of buffer) and centrifuging the homogenate at 10,000xg for 10 min. Absorbance readings (A405 nm) were recorded with a Metertech Σ 960 automatic ELISA plate reader. Three times the mean A405 nm readings from healthy samples was taken as the negative-positive threshold. At 30 dpi, positive samples were further analysed with anti-RYMV monoclonal antibodies A and D as described in Fargette et al. (2002) in order to determine the viral serotype therein.
For back inoculations tests, inoculums were prepared from leaf samples as described above. They were used to inoculate 2 week-old plantlets of the susceptible rice cultivar BG90-2 and symptom development was monitored for 30 dpi.
Identification of RYMV strains in inoculated plants: To determine if plant species selected specific RYMV strains from the mixture of isolates which was inoculated, two leaf samples of 0.5 g each were taken from five individual infected plants per host species. One of the samples was analysed by ELISA using monoclonal antibodies mab A and mab D as described by Fargette et al. (2002). The second sample was ground in phosphate buffer as indicated above and the extract was inoculated to two-week-old seedlings of rice cultivars Gigante and Tog 5681. Inoculated seedlings were kept for 45 dpi to monitor the development of symptoms.
Plant reaction to selected RYMV isolates: This experiment was conducted in order to ascertain the ability of host plants to differentiate between RYMV strains or pathogenic variants. A panel of virus isolates was used to inoculate five plants of every species identified as hosts of RYMV and of wild rice species O. barthii and O. longistaminata. The panel of RYMV isolates included isolates in Table 1 and four additional isolates (Ni 133, Td 6, Td 68 and Td 98) which belonged to serotype Ser1. Ni 133 and Td 6 were of pathotypes TG and G, respectively whereas Td 68 and Td 98 were both of pathotype S. Inoculations were done as indicated above and symptoms were monitored daily starting from the third day after inoculation.
RESULTS
Susceptibility of plant species to RYMV: In total 66 plant species including 40 Poaceae species, four other monocots and 22 dicotyledonous species were tested for susceptibility to RYMV (Table 2). At 12 to 21 days post-inoculation, symptoms were observed in seven species which belonged to two tribes of the monocotyledonous plant family Poaceae.
| Table 2: | Plant reaction to mechanical inoculation of RYMV |
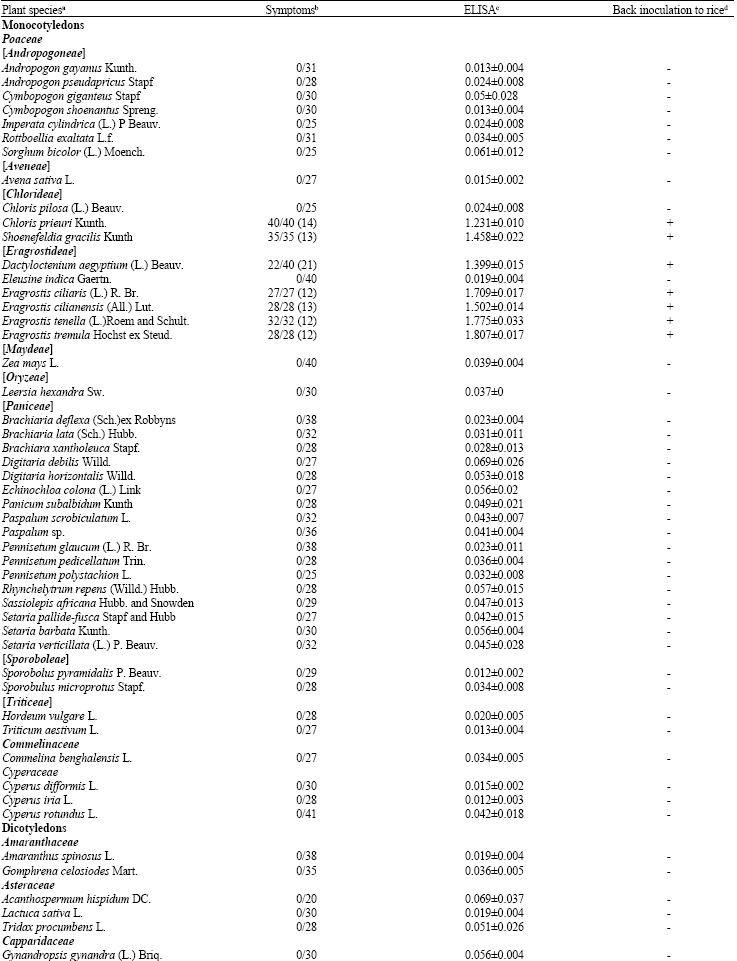 | |
 | |
aSubdivisions of the Poaceae family into tribes are indicated in square brackets; bNumber of symptom-bearing plants out of number inoculated. Mean virus incubation times in days post-inoculation are indicated in parentheses where applicable; cMean A405 nm±standard deviation; dPresence (+) or absence (-) of infection in the susceptible rice cultivar BG90-2 | |
The first tribe (Chorideae) contained only the two species Choris prieuri Kunth. and Shoenefeldia gracilis Kunth. whereas the second tribe (Eragrostideae) owned the five remaining species: Dactyloctenium aegyptium, Eragrostis ciliaris (L.) R. Br., E. cilianensis (All.) Lut., E. tenella (L.) Roem and Schult. and E. tremula Hochst. ex Steud. Of the seven species, C. prieuri, S. gracilis and E. cilianensis are newly recorded in the host range of RYMV. Symptoms induced consisted of systemic yellow or green mottling and mosaic on the leaf blades. In some species, particularly E. ciliaris, symptoms were very severe giving a whitish discoloration to the leaves. By contrast, the symptoms were less noticeable in C. prieuri plants and in some D. aegyptium plants. All symptomatic plants sustained reduced growth and some of them even die prematurely after 20 to 30 days post-inoculation. Like most grasses and other monocot species, dicotyledonous plants did not show any symptom. In particular, symptomless dicot species included several cultivars of the following crop plants: Zea mays L. (9 cultivars), Sorghum bicolor (L.) Moench. (7), Pennisetum glaucum (L.) R. Br. (4), Avena sativa L. (3), Triticum aestivum L. (3), Glycine max (L.) Merr. (5) and Sesamum indicum L. (3).
In serological analysis of leaf extracts, samples taken from symptomatic plants gave positive reactions confirming the presence of RYMV in the plants. For each species, one sample was taken from each of five plants chosen at random. Virus detection was unequivocal in all samples as A405 nm readings from diseased plants were 8 to 12 times greater than negative-positive cut-off values which did not exceed 0.15. Therefore, the seven Poaceae species in which plants symptoms were induced and RYMV detected are susceptible to the virus. For species where no symptom was induced, serological analyses were performed on samples taken from groups of five plants. As shown in Table 2, A405 nm readings obtained from these analyses were below the positive-negative thresholds indicating the absence of virus detection.
Back inoculations from inoculated plants of all species to susceptible rice BG90-2 were also successful only for symptomatic plants. Moreover, inoculums prepared from symptomatic plants were able to reproduce symptoms in healthy seedlings of the respective plants species. No symptom was induced from asymptomatic plants of any plant species, which confirmed the absence of virus detection in these plants.
Identification of RYMV strains in plant extracts: The use of anti-RYMV monoclonal antibodies (mab A and mab D) to test leaf samples from plants of the seven susceptible Poaceae species resulted in clear reaction of both antibodies with all samples except those from D. aegyptium. Extracts from D. aegyptium reacted with mab A only. The reaction profile obtained from most samples was characteristic of RYMV serotype Ser-Sa which reacts with both mab A and mab D. Therefore, the samples may have been infected by isolates of serotype Ser-Sa from the mixture of isolates inoculated.
| Table 3: | Reactions of wild host species to selected RYMV isolatesa |
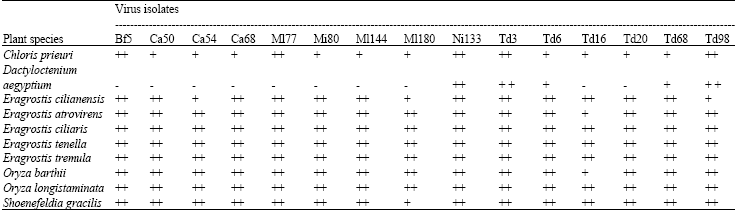 | |
| aFive plants were inoculated with each virus isolate and plant reactions were rated as follows: ++ (4-5 infected plants); + (2-3 plants infected) and - (no infection) | |
They could also have been co-infected by Ser1 and Ser2 isolates or by a mixture of one these two serotypes with isolates of Ser-Sa. At least, these results indicated that none of the susceptible plant species other than D. aegyptium was selectively infected by either Ser1 or Ser2. Reactions of D. aegyptium samples were typical of Ser1 isolates, indicating that isolates of other serotypes were unable to infect this plant species.
Inoculations of leaf extracts from the seven susceptible plant species to rice cultivars Gigante and Tog 5681 also resulted in a single pattern of pathogenic reaction. Extracts from all inoculated plants induced rice mottle symptoms in both rice cultivars, which was characteristic of pathotype TG. Therefore, leaf extracts contained isolates of pathotype TG or a mixture isolates of pathotypes T and G.
Reaction of plant species to specific RYMV strains: Except D. aegyptium, all other host species were infected by the 15 RYMV isolates inoculated (Table 3). Only five virus isolates were able to induce symptoms in D. aegyptium plants. No infection was obtained in this species by other isolates and absence of symptom expression was also confirmed by the lack of virus detection by ELISA (data not shown). Regardless of virus isolates, most species were highly susceptible as all inoculated plants were infected. However, infection rates were particularly low in Chloris prieuri and inoculated were plants rarely infected in total. Moreover, symptoms vanished progressively after the flowering stage although the plants remained ELISA positive. Inconspicuousness of symptoms also occurred in three plant species (D. aegyptium, E. tenella and S. gracilis) but this happened for only a few virus isolates.
DISCUSSION
Although sobemoviruses infect both monocot and dicot plants in more than 15 different families, host ranges of individual virus species are narrow (Tamm and Truve, 2000; Hull and Fargette, 2005). Plant species that have been reported to belong to RYMV host range are from the sole Poaceae family, which was confirmed by results obtained in this study. Despite the diversity of non-Gramineae species tested (26 members in 13 families) none of them was found to be susceptible to RYMV. Furthermore, Poaceae species tested belonged to nine tribes but it was shown that only two of them owned host plant species. In agreement with previous reports (Bakker, 1974; Bakker, 1975), most host species (five out of seven) belonged to the Eragrostidae tribe. Of these, Eragrostis cilianensis is reported for the first time. The two remaining host species (C. prieuri and S. gracilis) are also newly reported. Moreover, they are the first RYMV host species identified in the Chlorideae tribe. Apart from this tribe, RYMV host species have been found within six other tribes including Andropogoneae, Bromeae, Eragrostideae, Paniceae, Phalarideae and Oryzeae (Watson et al., 1985). Altogether, our results confirmed the narrowness of RYMV host range.
Like rice the main host, most plant species identified within RYMV host range showed clear mottling symptoms which were persistent. This is particularly useful for field identification of infected wild hosts. By contrast, inconspicuous symptoms observed in some D. aegyptium plants and in species such as C. prieuri and disappearance of symptoms in old plants of some species are tremendous obstacles for the identification of wild hosts. In such instances, infected plants need to be closely scrutinized. Surveying the plants at younger stages before flowering may ease the identification of those infected.
Disappearance of symptoms in old plants of species like E. tenella and in some plants of S. gracilis probably resulted from specific virus-host interactions. These symptomless infections differed from those obtained in E. tenella in East Africa where the plants never showed symptoms (Bakker, 1974). Specificity of interactions between RYMV and its host plants is clearly exemplified by D. aegyptium. Recently, this species was found to be susceptible to only one of four RYMV isolates tested (Allarangaye et al., 2006). Present results demonstrated that only five out of fifteen isolates were able to infect this species, which confirmed that D. aegyptium is a differential host plant for RYMV. However, the mechanism by which this species is infected or not by virus isolates remains unexplained. Indeed, all five isolates which infected D. aegyptium plants belonged to serotype Ser1 but they were of different pathotypes. Moreover, some Ser1 isolates e.g., Bf 5, Ca 50, Td 16 or Td 20 were unable to infect D. aegyptium (Table 3).
Specific isolate-host interactions may also be responsible for contrasting results observed with some plant species about their status of being or not host plants for RYMV. For example, our results identified Eragrostis cilianensis as a systemic host species whereas this species was found to be insusceptible to RYMV elsewhere (Bakker, 1974). Similarly, species such as Digitaria sanguinalis (L.) Scop, Echinochloa crus-galli (L.) P.B., Eragrostis atrovirens, E. tenuifolia (A. Rich.) Steud, Panicum maximum Jacq and Panicum repens identified as experimental or natural host species in West Africa (Awoderu, 1991; Konaté et al., 1997; Traoré, 2006) were not infected by the virus in East Africa (Bakker, 1974). Such discrepancies seemed to be not geography-based. Within West Africa, the wild finger-millet Eleusine indica Gaertn. was reported as highly susceptible to RYMV (Awoderu, 1991) in the contrary of our results and also those of Bakker in East Africa.
Possibly, differences observed in RYMV host range studies could be bypassed by using a diversity of isolates in the same inoculum. Our results indicated that host plant species showed unique patterns of reaction upon inoculation of the mixture of isolates. Yet, they were found susceptible to individual RYMV isolates which composed the mixture (Table 3). Consequently, interactions between virus isolates in the mixture, if any, did not prevent simultaneous infections by individual isolates. Therefore, the use of a mixture of isolates to identify RYMV host species appeared to be more appropriate than the lengthier and more tiresome test of individual virus isolates.
Overall, in addition of being narrow, RYMV host range is complex. In the one hand, RYMV host species belong to highly heterogeneous Poaceae groups. The seven tribes in which host species have been identified are from four grass subfamilies (Chloridoideae, Ehrhartoideae = Oryzoideae, Panicoideae and Pooideae) with distinct phylogenies. Chloridoideae and Panicoideae are part of the same lineage while Ehrhartoideae and Pooideae are members of another lineage (Kellogg, 2000; Hilu, 2004). In the other hand, plant species found to be insusceptible to RYMV often belonged to tribes or even genus which contained host species. A clear example is given by members of genus Oryza and Leersia within the Oryzeae tribe. The genus Oryza is composed of well-known host species such as Oryza sativa, O. barthii, O. glaberrima and O. longistaminata but members of the tribe Leersia were found to be insusceptible to RYMV (Bakker, 1974; this study). At the genus level, like several members of the genus Chloris including Chloris gayana Kunth., Chloris pycnothrix Trin., Chloris roxburghiana Schult. (Bakker, 1974), we were unable to infect Chloris pilosa Sch and Thonn. whereas Chloris prieuri was identified as a host species for RYMV.
ACKNOWLEDGMENTS
This study was supported by a fellowship of the French Government through the French Ministry for Cooperation to M.D. Allarangaye. Technical assistance of Mr Zerbo Doumacé is greatly acknowledged.
REFERENCES
- Abo, M.E., M.D. Alegbejo, A.A. Sy and S.M. Misari, 2000. An overview of the mode of transmission, host plants and methods of detection of Rice yellow mottle virus. J. Sustain. Agric., 17: 19-36.
CrossRefDirect Link - Allarangaye, M.D., O. Traore, E.V.S. Traore, R.J. Millogo and G. Konate, 2006. Evidence of non-transmission of Rice yellow mottle virusthrough seeds of wild host species. J. Plant Pathol., 88: 307-313.
Direct Link - Awoderu, V.A., 1991. Rice yellow mottle virus in West Africa. Trop. Pest Manage., 37: 356-362.
Direct Link - Bakker, W., 1970. Rice yellow mottle, a mechanically transmissible virus disease of rice in Kenya. Neth. J. Plant Pathol., 76: 53-63.
CrossRefDirect Link - Banwo, O.O. and R.H. Makundi, 2001. Newly recorded species of Chaetocnema, vector of Rice yellow mottle virus in Tanzania. N. Z. J. Crop Hortic. Sci., 29: 61-65.
CrossRefDirect Link - Fargette, D., A. Pinel, H. Halimi, C. Brugidou, C. Fauquet and M. van Regenmortel, 2002. Comparison of molecular and immunological typing of the isolates of rice yellow mottle virus. Arch. Virol., 147: 583-596.
CrossRefDirect Link - Fargette, D., A. Pinel, Z. Abubakar, O.Z. Traore and C. Brugidou et al., 2004. Inferring the evolutionary history of rice yellow mottle virus from genomic, phylogenetic and phylogeographic studies. J. Virol., 78: 3252-3261.
CrossRefDirect Link - Hilu, K.W., 2004. Phylogenetics and chromosomal evolution in the Poaceae (grasses). Aust. J. Bot., 52: 13-22.
CrossRefDirect Link - Konate, G., O. Traore and M.M. Coulibaly, 1997. Characterization of Rice yellow mottle virusisolates in sudano-sahelian areas. Arch. Virol., 142: 1117-1124.
Direct Link - Konate, G., S. Sarra and O. Traore, 2001. Rice yellow mottle is seed-borne but not seed transmitted in rice seeds. Eur. J. Plant Pathol., 107: 361-364.
CrossRefDirect Link - Pinel, A., P. N'Guessan, M. Bousalem and D. Fargette, 2000. Molecular variability of geographically distinct isolates of rice yellow mottle virus in Africa. Arch. Virol., 145: 1621-1638.
CrossRefDirect Link - Sarra, S. and D. Peters, 2003. Rice yellow mottle virus is transmitted by cows, donkeys and grass rats in irrigated rice crops. Plant Dis., 87: 804-808.
Direct Link - Sarra, S., P. Oevering, S. Guindo and D. Peters, 2004. Wind-mediated spread of Rice yellow mottle virus (RYMV) in irrigated rice crops. Plant Pathol., 53: 148-153.
Direct Link - Traore, O., F. Sorho, A. Pinel, Z. Abubakar and O. Banwo et al., 2005. Processes of diversification and dispersion of rice yellow mottle virus inferred from large-scale and high-resolution phylogeographical studies. Mol. Ecol., 14: 2097-2110.
CrossRefDirect Link - Traore, O., A. Pinel, E. Hebrard, M.Y.D. Gumedzoe, D. Fargette, A.S. Traore and G. Konate, 2006. Occurrence of resistance-breaking isolates of rice yellow mottle virus in West and central Africa. Plant Dis., 90: 259-263.
Direct Link - Traore, O., M. Traore D. Fargette, and G. Konate, 2006. Rice seedbeds as a source of primary infection by rice yellow mottle virus. Eur. J. Plant Pathol., 115: 181-186.
Direct Link