
ismet Yildirm
Department of Plant Protection, Faculty of Agriculture,Çanakkale Onsekiz Mart University,Terzioglu Campus, 17020 Çanakkale, Turkey
Binnur Meriçli Yapici
Department of Biology, Faculty of Arts and Sciences, Çanakkale Onsekiz Mart University,Terzioglu Campus, 17020 Çanakkale, Turkey
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 8 | Page No.: 1294-1300







ABSTRACT
Fungal activities of food additives, potassium sorbate, methylparaben, sodium benzoate, propylparaben, sorbic acid and plant activators harpin-protein and potassium dioxide were comparatively examined with ipradion in vitro conditions on two isolates of Botrytis cinerea obtained from strawberry fruits. Food additives and plant activators showed inhibitory effect at different levels on the mycelial growth of fungus isolates. According to the Minimum Inhibition Concentration (MIC), sorbic acid (300 μg mL-1 for each isolate) and comparison fungicide ipradion (10 μg mL-1 for each isolate) displayed the highest inhibition effect on the radial mycelial growth of fungal isolates. Similar efficacy was obtained at the highest dose (1000 μg mL-1) from other additives propylparaben (for both isolates), potassium sorbet and methylparaben (for isolate 1) and from plant activator potassium oxide. It was detected that all alternative substances has high inhibition activity according to ED50 values. Methylparaben, harpin-protein and potassium oxide revealed efficacies similar to ipradion. While harpin-protein did not have any effect on B. Cinerea germination, other alternative substances excluding potassium sorbate inhibited germination completely at differing doses. Potassium oxide and food additives except potassium sorbate inhibited the spore germination of isolates by 50% at doses between 25-345 μg mL-1. MIC value of ipradion on spore germination of isolate-1 and isolate-2 was determined as 10 and 30 μg mL-1, while ED50 values were determined as 2.9 and 12.5 μg mL-1, respectively. Data obtained from the research verified that food additives may be used in traditional and organic agriculture individually, or in appropriate combination with each other or with fungicides. Moreover, the study put forth that harpin-protein and potassium oxide not only activate plants against pathogens, but they may also have toxic effect on different growth periods of fungus. Although ipradion works only with two isolates, its MIC and ED50 values were considered noteworthy in terms of the resistance risk of B. cinerea.
PDF Abstract XML References Citation
How to cite this article
ismet Yildirm and Binnur Meriçli Yapici, 2007. Inhibition of Conidia Germination and Mycelial Growth of Botrytis cinerea by Some Alternative Chemicals. Pakistan Journal of Biological Sciences, 10: 1294-1300.
DOI: 10.3923/pjbs.2007.1294.1300
URL: https://scialert.net/abstract/?doi=pjbs.2007.1294.1300
DOI: 10.3923/pjbs.2007.1294.1300
URL: https://scialert.net/abstract/?doi=pjbs.2007.1294.1300
INTRODUCTION
Botrytis cinerea Pers: Fr. is an important pathogen that has a broad host range including vegetables, indoor plants, bulbed plants and fruits. If grey mould disease (B. cinerea) cannot be prevented adequately, it may cause important losses in efficiency and quality (Coley-Smith et al., 1980; Erkan et al., 1997; Body-Wilson et al., 1998; Baraldi, 2002). The disease may be controlled in classical fungicides; however, most of the traditional fungicides embody the risk of leaving remnants on products and have negative effects on human beings, animals and environment (Erkan et al., 1997). Furthermore, B. cinerea may easily develop races with high resistance against many synthetic fungicides (AbouJawdah and Itani, 1994; Erkan et al., 1997; Knight et al., 1997; Palmer et al., 1997; Vallejo et al., 2003). On these grounds, recent studies have focused on alternative chemicals and biologic agents in the control of pathogens (Antonov et al., 1997; Archbold et al., 1997; Boyd-Wilson et al., 1998; Reddy et al., 1998; Ntirampemba, 1998). Plant extracts, sodium and potassium salts and some antimicrobial food additives have gained importance in the control of pathogens.
In this study, the effects of food additives, potassium sorbate, sodium benzoate, methylparaben, propylparaben, sorbic acid and potassium dioxide and harpin-protein in vitro on the mycelium growth and conidia germination of B. cinerea were determined in vitro.
MATERIALS AND METHODS
Origin of B. cinerea isolates: B. cinerea isolates (I1, I2) were isolated from infected fresh strawberry fruits (C1; Camorosa ve SC2; Sivic Charli) obtained from a greenhouse in Aydın (Turkey).
| Table 1: | Some properties of the test chemicals |
 | |
Isolates were developed in an incubator at 21°C in PDA environment and were transferred into tubes with PDA in order to obtain stock cultures.
Natural chemicals: Potassium-sorbate, methylparaben (methyl-p-hydroxylbenzoate), harpin-protein, potassium dioxide, sodium benzoate, propylparaben (propyl-p-hydroxybenzoate), sorbic acid and ipradion as comparison fungicide were tested in vitro so as to determine the efficiency of alternative chemicals against B. cinerea. Test chemicals were used at doses of 0 (control), 0.3, 1, 3, 10, 30, 100, 300 and 1000 μg mL-1. In the experiments, streptomycin sulfate (0.3 μg mL-1) was added to the culture environment (Delen et al., 1984). Some properties of test chemicals and comparison fungicides are given in Table 1.
Efficiency of test chemicals on mycelium growth: Stock isolates were developed on PDA environment and agar discs (Ø4 mm) cut from fungal culture using a sterile cork borer were planted on the surface of MM environment with and without test chemicals by placing the side with fungal culture against the bottom. After inoculation, petri dishes wrapped with parafilm were incubated at 25°C in the dark. Colony diameters were measured at the widest point for 3 days at 24 h intervals. Inhibitory effects of chemicals were detected by comparing the diameters of the control group with the diameters of colonies developed in nutrition environment treated with chemicals. ED50 values were found using semi-logarithmic graphics. The trials were established three repetition and coincidence blocks trail design.
Inhibition of conidia germination: Efficiency of test chemicals on the conidia germination of B. cinerea was tested in MM environment and it was compared with the efficiency of ipradion. For this purpose, isolates were transferred from stock cultures to MM environment and they were incubated in the incubators for 10 days at 21°C in the dark for 12 h and in the light for 12 h alternately. Later, MM environments with and without test chemicals were inoculated in a glass tower (0.35x0.70 m2 high) with conidia obtained from fungus cultures (30 conidia mm-2). Conidia obtained from cultures were given to the environments by means of an air current provided by an air pump. Inoculated petry dishes were incubated at an optimum temperature of 20°C in the dark for the germination of B. cinerea conidia. In order to stop conidial growth at the end of 24 h after inoculation, blotting papers pasted inside petry stoppers were treated with formaldehyde (7%). Conidia germination percentage was determined by observing 200 spores under microscope for each recurrence. Conidia germination was verified if the length of germ tube was equal to or more than the spore (Vicedo et al., 2006). Treatment was established four repetition and coincidence blocks trial design. Inhibitory effects of chemicals were determined by means of Abbot Formula and ED50 values were found using semi-logarithmic graphics.
RESULTS
Efficacies of chemicals on colonial growth of B. cinerea (isolate-1) are given in Fig. 1 and 2. Colonial growth of both isolates was generally inhibited to a significant extent even in the lowest doses of test chemicals (Fig. 1 and 2).
It was detected that ipradion at low doses (0.3-3 μg mL-1) with a minimum 3% and maximum 100% inhibition rate is the most effective chemical in inhibiting colonial growth and it is followed by methylparaben, bioclean, harpin-protein and others respectively (Fig. 1). Sorbic acid at 1000 μg mL-1 dose reached the highest efficacy (100%) on colonial growth of isolate-1. On the other h and, harpin-protein at 1000 μg mL-1 dose revealed the lowest efficacy (74.7%) among test chemicals.
Efficacies of test chemicals on the colonial growth of isolate-2 were similar to the efficacies of that of isolate-1. Ipradion, methylparaben, harpin-protein and potassium oxide at low doses displayed important efficacies on colonial growth of isolate-2; however, other chemicals were effective on colonial growth only at doses over 30 μg mL-1 (Fig. 2).
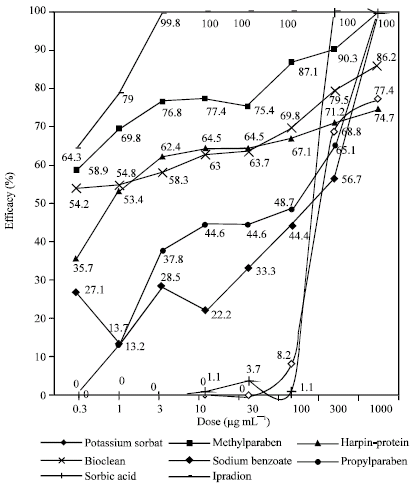 | |
| Fig. 1: | Effects of the chemicals on colonial growth of Botrytis cinerea (Isolate-1) |
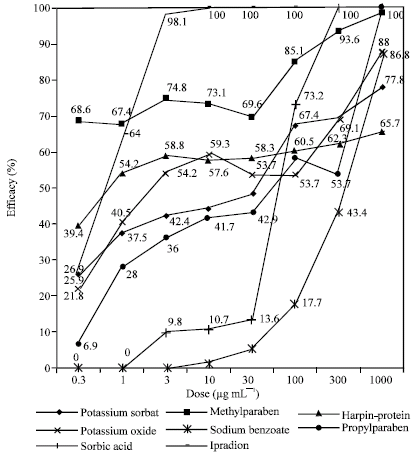 | |
| Fig. 2: | Effects of the chemicals on colonial growth of B. cinerea (Isolate-2) |
| Table 2: | MIC and ED50 values of test chemicals on mycelial growth of Botrytis cinerea |
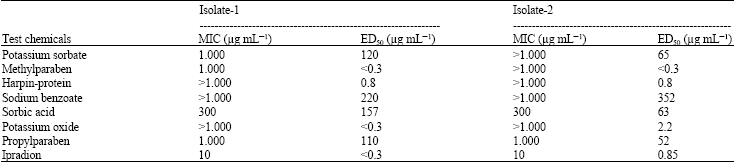 | |
| Table 3: | Efficacies of chemicals on Conidia germination of B. cinerea (Isolate-1) (%) |
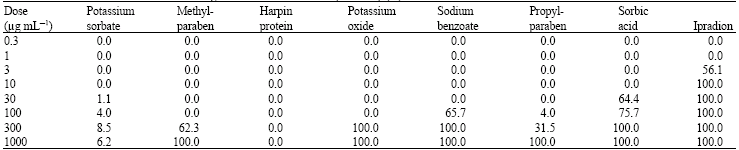 | |
| Table 4: | Efficacies of chemicals on Conidia germination of B. cinerea (Isolate-2) (%) |
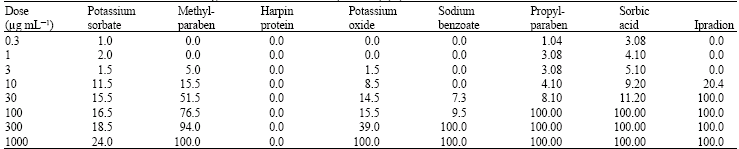 | |
Minimum Inhibition Concentrations (MIC) and ED50 values of test chemicals on radial mycelial growth of B. cinerea are introduced in Table 2.
Ipradion (comparison fungicide) at a dose of 10 μg mL-1 completely inhibited the radial mycelial growth of both isolates of B. cinerea. It was followed by sorbic acid at a dose of 300 μg mL-1. MIC values of other alternative chemicals were detected at 1000 μg mL-1 and higher doses (Table 2). On the other h and, ED50 values of methylparaben, potassium oxide and harpin-protein were close to the values of Ipradion. ED50 values of other alternative chemicals were slightly higher than that of Ipradion (Table 2). Ipradion at a dose of 3 μg mL-1 reduced conidia germination of B. cinerea (Isolate-1) to a significant extent, whereas alternative chemicals at low doses failed to inhibit conidia germination of B. cinerea (Table 3). Sorbic acid at a concentration of 30 μg mL-1 inhibited conidia germination considerably and it was followed by sodium benzoate at 100 μg mL-1 dose, potassium oxide at 300 μg mL-1 dose and methylparaben respectively. On the other h and, while harpin-protein did not show any efficacy on conidia germination of B. cinerea (isolate-1), potassium sorbate could not reveal sufficient efficacy on conidia germination of B. cinerea. Potassium oxide, sorbic acid and sodium benzoate at 300 μg mL-1 completely inhibited conidia germination (Table 3).
While comparison fungicide Ipradion inhibited conidia germination of Isolate-2 to 20.4% at a dose of 10 μg mL-1, it had a complete inhibitory effect at 30 μg mL-1 and higher doses (Table 4). It was determined that sorbic acid and propylparaben at 100 μg mL-1 dose, among alternative substances, have the lowest minimum inhibition concentration on conidia germination of Isolate-2 and these are followed by sodium benzoate (300 μg mL-1), methylparaben (1000 μg mL-1) and potassium oxide (1000 μg mL-1) respectively. While harpin-protein did not have any efficacy on conidia germination of isolate-2, MIC value of potassium sorbate was detected to be above 1000 μg mL-1 (Table 4).
ED50 values of test chemicals on conidial germination of B. cinerea isolates are introduced in Table 5.
Among test chemicals, ipradion at doses of 2.9 and 12.5 μg mL-1 had the lowest ED50 values on conidial germination of both isolates and it was followed by sorbic acid (Table 5). ED50 values of methylparaben, propylparaben, sodium benzoate and potassium oxide revealed differences depending on the isolates.
| Table 5: | ED50’s of test chemicals on Conidial germination of Botrytis cinerea |
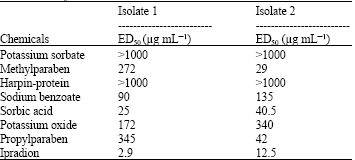 | |
ED50 values of potassium sorbate and harpin-protein were determined to be 1000 μg mL-1 higher for both isolates.
DISCUSSION
In this study; potassium dioxide and harpin protein besides potassium sorbate, sodium benzoate, methylparaben, propylparaben and sorbic acid, which are food additives that may be alternatives to classical fungicides, were analysed in vitro conditions in comparison with a classic fungicide ipradion regarding their efficacies on mycelium growth and conidia germination of Botrytis cinerea.
Among alternative substances, food additives potassium sorbate, sodium benzoate, methylparaben, propylparaben and sorbic acid and harpin-protein and potassium oxide showed strong inhibition effect on mycelial growth and conidia germination of B. cinerea at increasing doses. Food additives are being used widely for a long time in cosmetics and in the field of medicine, as well as in food preservation due to their anti-microbial effects and they have high efficacy against yeast and mould (Wade and Weller, 1994). Effect mechanisms of sorbic acids, benzoic acids and parabens against yeast and fungi rely on two factors: decreasing of pH in the intracellular part because of becoming ionized of fungi’s non-decomposing acid molecules or damaging of substrate transporting due to changing of cell-membrane’s permeability (Liewen, 1991; Khan et al., 2001; Vicedo et al., 2006). Enzyme activation in fungi depends on pH value and performance of extracellular enzyme catalysis decreases at pH levels that are not within optimum levels (Dix and Webster, 1995). Therefore, fungus cannot provide the sufficient energy for hif development (Skirdal and Eklund, 1993).
It was verified in the study that especially sorbic acid inhibits mycelial growth of both isolates (isolate-1, isolate-2) of B. cinerea at low doses. Similarly, it was noted that different forms of sorbic acid play a considerable role in the inhibition of fungal growth of Penicillium chrysogenum, Cladosporium cladosporioides and Ulocladium atrum (Palmer et al., 1997). Sorbic acid at doses of 100 and 300 μg mL-1 completely inhibited conidia of both isolates of B. cinerea and proved to be successful in the inhibition of mycelial growth. It was also detected that sorbic acid has low ED50 values, such as 25 μg mL-1 (for isolate-1) and 40.5 μg mL-1 (for isolate-2), besides low MIC values on chemical germination. Although sorbic acid effectively inhibits radial mycelia growth and spore germination of B. cinerea, its hydrophobic structure makes it difficult to use it in practice as antifungal agent. Dudman (1963) notes that using water-soluble salts of sorbic acid, such as sorbate and sodium sorbate, may provide a solution in practice. Although potassium sorbate in this study could not show sufficient efficacy on conidia germination (ED50 and MIC values for both isolates were higher than 1000 μg μg mL-1), it inhibited mycelial growth of fungus to a great extent. When the significance of mycelial growth in disease development is taken into consideration, potassium sorbate may be regarded as successful.
Other additives, parabens (methylparaben, propylparaben) and sodium benzoate, showed high inhibitory effect on radial mycelia growth and conidia germination of both isolates (isolate-1 and isolate-2) of the pathogen. Sodium benzoates and parabens are used in additives to inhibit fungal mould (Khan et al., 2001).
Sodium benzoate could not totally inhibit mycelia of isolates in vitro, but ED50 values were found to be at a low level (220 μg mL-1 for isolate-1; 352 μg mL-1 for isolate-2). Sodium benzoate displayed a higher efficacy on conidia germination of B. cinerea isolates. Clausen and Yang (2003) noted that sodium benzoate and potassium sorbate at doses of 5% inhibited the spores of Penicillium chrysogenum, Aspergillus niger and Trichoderma viride and also stated that sodium nitrate at dose combinations of 0.3%, in addition to especially these two chemicals, totally inhibited the spores of all three fungi in malt agar environment.
Other additives, methylparaben and propylparaben, had important inhibitory effects on radial mycelia growth and conidia germination of B. cinerea isolates. It was observed that efficacy of methylparaben on mycelia growth of fungus was close to that of comparison fungicide ipradion. While LD50 value of the chemical in isolate-1 (ED50<0.3 μg mL-1) was the same with the value of ipradion, it was noteworthy that this value was at lower doses (ED50<0.3 μg mL-1) than ipradion (ED50 = 0.85 μg mL-1) in isolate-2. However, ED50 values on spore germination were observed to be slightly higher than ipradion in both isolates. MIC values of the chemical were determined to be 1000 μg mL-1 on mycelia growth and spore germination of both isolates. In a study carried out with mould fungi, it was observed that methylparaben displayed different MIC values depending on fungus type; for example, 600 μg mL-1 against Aspergillus oryzae, 1000 μg mL-1 against Aspergillus niger and 250 μg mL-1 against Trichoderma lignorum (Haag and Loncrini, 1984).
It was observed that propylparaben at low doses had a higher inhibitory effect on mycelia growth of B. cinerea isolates than the effect of sorbic acid and sodium benzoate. Yet, MIC value determined as 1000 μg mL-1 against both isolates was higher than sorbic acid. Propylparaben completely inhibited conidia germination of isolate-1 and isolate-2 at doses of 1000 and 100 μg mL-1, respectively. ED50 values on germinations were similar to the values of methylparaben.
Apart from the food additives included in the study, potassium oxide and harpin-protein displayed a medium level of inhibition effect on mycelia growth of fungus. While ED50 values of both substances on mycelia growth were detected to be quite low, their MIC values were above the dose of 1000 μg mL-1. Harpin-protein did not have any efficacy on conidia germination of B. cinerea isolates, whereas potassium oxide showed efficacy only at high doses. ED50 value of harpin-protein was above 1000 μg mL-1 for both isolates as observed in mycelia growth, but the values of potassium oxide were detected to be lower. Although it is known that harpin-protein is not directly effective on microorganisms, but stimulates resistance against pathogens in plants (EPA, 2006); it was considered noteworthy that it inhibited especially mycelia growth of fungus in this study. This result indicates that harpin-protein can have direct toxic effect on mycelia of fungi.
Similar to harpin-protein, potassium oxide also strengthens the defence mechanism of plants rather than providing a direct effect on the pathogens. The efficacy of potassium oxide on fungal growth may be considered as a direct toxic effect.
In the study, comparison fungicide ipradion revealed high inhibitory effect both on mycelia growth and conidia germination of B. cinerea. Erkan et al. (1997) determined in their study, in which they examined the sensitivity of B. cinerea isolates obtained from vineyards towards dicarboximide fungicides, that isolates have high sensitivity towards ipradion and their ED50 values on mycelia growth of fungus and MIC value on spore germination are both lower than 0.3-1.7 μg mL-1. However, it is significant that ED50 values obtained on spore germination (2.9 μg mL-1 for isolate-1; 12.5 μg mL-1 for isolate-2) indicate that isolates are sensitive to ipradion at low and medium levels (Erkan et al., 1997).
Data obtained from the study have proved that food additives in vitro inhibit mycelia growth and spore germination of B. cinerea to a significant extent especially at high doses. This shows that food additives, alone or in combinations, may replace classical fungicides in organic and traditional agriculture in practice. Furthermore, synthetic fungicides may also be used in combination with food additives in order to increase their efficacies. Yet, in the first place, it requires researches to be carried out on plants. Subsequently, it has been determined in this study that harpin-protein and potassium oxide not only activate plants against disease factors in practice, but also have toxic effects on pathogens.
REFERENCES
- Archbold, D.D., T.R. HamiltonKemp, M.M. Barth and B.E. Langlois, 1997. Identifying natural volatile compounds that control gray mold (Botrytis cinerea) during postharvest storage of strawberry, blackberry and grape. J. Agric. Food Chem., 45: 4032-4037.
CrossRefDirect Link - Baraldi, E., P. Bertolini, E. Chierici, B. Trufelli and D. Luiselli, 2002. Genetic diversity between Botrytis cinerea isolates from unstored and cold stored Kiwi fruit. J. Phythopathal., 150: 629-635.
CrossRefDirect Link - Dudman, W.F., 1963. Sorbic hydroxamic acid, an antifungal agent effective over a wide pH range. Applied Microbiol., 11: 362-364.
Direct Link - Khan, S.H., J. Aked and N. Magan, 2001. Control of the anthracnose pathogen of banana (Colletotrichum musae) using antioxidants alone and in combination with thiabendazole or imazalil. Plant Pathol., 50: 601-608.
Direct Link - Knight, S.C., V.M. Anthony, A.M. Brady, A.J. Greenland and S.P. Heaney et al., 1997. Rationale and perspectives on the development of fungicides. Annu. Rev. Phytopathol., 35: 349-372.
CrossRefDirect Link - Ntirampemba, G., B.E. Langlois, D.D. Archbold, T.R. Hamilton-Kemp and M.M. Barth, 1998. Microbial populations of Botrytis cinerea-inoculated strawberry fruit exposed to four volatile compounds. J. Food Protect., 61: 1352-1357.
Direct Link - Palmer, C.L., R.K. Horst and R.W. Langhans, 1997. Use of bicarbonates to inhibit in vitro colony growth of Botrytis cinerea. Plant Dis., 81: 1432-1438.
CrossRefDirect Link - Reddy, M.V.B., P. Angers, A. Gosselin and J. Arul, 1998. Characterization and use of essential oil from Thymus vulgaris against Botrytis cinerea and Rhizopus stolonifer in strawberry fruits. Phytochemistry, 47: 1515-1520.
CrossRefDirect Link - Skirdal, I.M. and T. Eklund, 1993. Microculture model studies on the effect of sorbic acid on Penicillium chrysogenum, Cladosporium cladosporioides and Ulocladium atrum at different pH levels. J. Applied Bacteriol., 74: 191-195.
CrossRefDirect Link - Vallejo, I., F. Munoz, M. Carbu, L. Rebordinos, F.J. Fernandez-Acero and J.M. Cantrol, 2003. Study on fungicide resistance of Botrytis cinerea isolates from diseased strawberry plants. Arch. Phytopathol. Plant Prot., 36: 1-7.
CrossRefDirect Link - Vicedo, B., M.O. Leyva, V. Flors, I. Finiti and G.A.D. Walters et al., 2006. Control of the phytopathogen Botrytis cinerea using adipic acid monoethyl ester. Arch. Microbiol., 184: 316-326.
Direct Link