
Yasar Durmaz
Department of Aquaculture, Fisheries Faculty, Ege University, 35100 Bornova Izmir, Turkey
Margarida Monteiro
Department Inovação Tecnológica e Valorização dos Produtos da Pesca,Instituto de Investigação das Pescas e do Mar-IPIMAR. Av. Brasília, 1449-006, Lisboa, Portugal
Edis Koru
Department of Aquaculture, Fisheries Faculty, Ege University, 35100 Bornova Izmir, Turkey
Narcisa Bandarra
Department Inovação Tecnológica e Valorização dos Produtos da Pesca,Instituto de Investigação das Pescas e do Mar-IPIMAR. Av. Brasília, 1449-006, Lisboa, Portugal
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 7 | Page No.: 1144-1146







ABSTRACT
Abstract: The objective of this study was to investigate sterols content of Porphyridium cruentum batch cultured in laboratory at 18° C and harvested in the stationary. The sterol distribution of this species is characterized by a predominance of cholesterol, with values as 199.0 mg 100 g- 1 freeze dry weight (92.2%). The second most important sterol was stigmasterol (4.9%) followed by β -sitosterol (2.2%). Studied sterols give to this species a special importance in for being used in food as supplements/nutraceuticals (including aquaculture).
PDF Abstract XML References Citation
How to cite this article
Yasar Durmaz, Margarida Monteiro, Edis Koru and Narcisa Bandarra, 2007. Concentration of Sterols of Porphyridium cruentum Biomass at Stationary Phase. Pakistan Journal of Biological Sciences, 10: 1144-1146.
DOI: 10.3923/pjbs.2007.1144.1146
URL: https://scialert.net/abstract/?doi=pjbs.2007.1144.1146
DOI: 10.3923/pjbs.2007.1144.1146
URL: https://scialert.net/abstract/?doi=pjbs.2007.1144.1146
INTRODUCTION
The microalgae are valuable natural products used as animal and human food sources. Important bioactive metabolites such as sterols (Volkman et al., 1981). Sterols, as chemical constituents of microalgae, are of increasing interest since: (1) the presence of these natural products in microalgae determines their food values; (2) these compounds are useful biomarkers for identifying sources of organic matter in sediments (Ponomarenko et al., 2004) and (3) they are not only essential components of biomembranes, but function in cell proliferation and signal transduction of microalga and other eukariotic organisms, modulating the activity of membrane-bound enzymes (Hartmann, 1998; Pioronen et al., 2000; Volkman, 2003; Ponomarenko et al., 2004).
The unicellular red alga Porphyridium cruentum (Rhodophyta, Porphyridiales) can be found in sea water and humid soils. The 4 and 9 μm diameter spherical P. cruentum cells lack a cell-wall and the cells can be solitary or massed together into irregular colonies held in mucilage (Vonshak, 1988). Microalgal cell metabolites which are mainly produced in the stationary growth phase are called secondary metabolites (Schelegel, 1985) and can include organic acids, carbohydrates, amino acids and peptides, vitamins, growth substances, antibiotics, enzymes and toxic compounds (Vonshak, 1986; Becker, 1994; Fuentes et al., 2000). The biochemical composition of P. cruentum shows that it is rich in many important compounds, i.e. the protein content of P. cruentum ranges from 28 to 39%, the carbohydrate content from 40 and 57% and the total lipids may reach 9-14% of dry weight and its biomass contains tocopherol, vitamin K and large amounts of carotenes (Becker, 1994).
The objective of this study was determined to concentration of sterols of P. cruentum, in batch cultures. On the other hand, this knowledge is also important for marine aquaculture programs.
MATERIALS AND METHODS
Microrganism: Porphyridium cruentum was obtained from the Microalgal Biotechnology Laboratory at Ben-Gurion University, Israel.
Culture conditions: The cells were grown in the laboratory in 10 L batch cultures. The alga was cultured in enriched artificial seawater (2.8 g L-1 NaCl) in Porphyridium medium (Vonshak, 1988). Cultures were kept at 18±1°C constant temperature room and continuous illumination was made with fluorescent lamps (Philips TLM 40W/54RS), at irradiance level of 196 μmol m-2 s-1 (Li-Core 195). When the culture reached 3.74x107 cells mL-1 at day 20, half of culture was harvested for analysis. Harvesting of the microalgae was done by flocculation with FeCl3, followed by centrifugation as reported by Batista and Martins (1991). The pellet obtained was freeze-dried for further analysis.
Sterols: The extraction of sterols was started with a saponification step mixing 0.25 g of freeze-dried material with 2.5 mL of 2M methanolic KOH solution and 100 μ L of 0.5% α -cholestanol in chloroform was added as internal standard. The mixture was placed in a water bath at 80° C during 30 min. After cooling 2.5 mL of cyclohexane and 2.5 mL of distilled water were added. The tubes were swirled for 1 min and centrifuged (Sigma 2K15 of B Braun, Germany) at 2150 g for 5 min. The organic phase was recovered, filtered and dried with anhydrous sodium sulphate. The extraction with cyclohexane was repeated twice but no water was added. The cyclohexane of the combined organic phases was evaporated and the residue obtained dissolved in 1 mL of n-hexane. This fraction was applied in a solid phase extraction column with 6 mL volume and 0.5 g of silica from Isolute-Spe (United Kingdom) in order to isolate the sterolic fraction. The column was previously conditioned with 5 mL of n-hexane. The sample (1 mL) was then applied and eluted with 5 mL of n-hexane. A second elution with 5 mL of a mixture of n-hexane: ethyl ether (9:1 v/v) was done and both eluates were discarded. A third elution with 5 mL of a mixture n-hexane: ethyl ether (1:1 v/v) was performed and the eluate recovered (the sterolic fraction). The solvent was evaporated and the obtained residue dissolved in 0.5 mL of chloroform. For the determination of the different sterols a previous silylation was done. The derivatization was performed with 0.1 mL of the mixture (99:1) BSTFA:TMCS (BSTFA-bis(trimethylsilyl)-trifluoracetamid and TMCS-trimethylchlorosilan from Fluka (St. Louis, Germany) and 0.1 mL of anhydrous pyridine Seccosolv from Merck (Germany) at 60° C during 30 min.
The solvent was eliminated under nitrogen and the residue dissolved in 0.1 mL of n-hexane. The trimethylsilylethers were analysed in a gas chromatograph Hewlett Packard HP 5890 A Series (USA) equipped with a flame ionisation detector at 300°C with manual injection in a split injector (100:1 split ratio) at 285°C. The separation was carried out with helium at a flow of 1 mL/min as carrier gas in a capillary column HP-5 (30 mx0.25 mm i.d.x0.25 μm film thickness) (USA) programmed for a constant temperature of 280°C.
RESULTS AND DISCUSSION
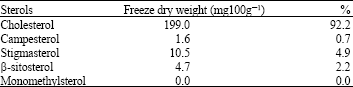
Sterols composition of P. cruentum cultured at 18° C and harvested in the stationary phase is shown in Table 1. Four sterol composition of P. cruentum were identified: cholesterol, campesterol, stigmasterol, β -sitosterol and one non-identified monomethylsterol. The sterol distribution of this species is characterized by a predominance of cholesterol, with values as 199.0 mg 100 g- 1 freeze dry weight (92.2%). The second most important sterol was stigmasterol (4.9%) followed by β -sitosterol (2.2%).
| Table 1: | Sterols (mg 100 g- 1 freeze dry weight) content of P. cruentum. The cultures were harvested at 20 days in the stationary phase, when cultures were harvested for the determination of sterols |
 | |
Many different sterol patterns have been found in microalgae and some of these are restricted to a particular class of microalgae. For example, diatoms display both simple and complex distributions often dominated by cholesterol (Volkman, 1986; Barrett et al., 1995). Many dinoflagellates contain mixtures of 10 or more 4-desmethyl and 4-methylsterols (Withers, 1983).
Soudant et al. (1998) had identified the epibrassicasterol or its isomer brassicasterol (96%) as the predominant sterol of T-Isochrysis, Pavlova lutheri showed a larger diversity in sterols with β-sitosterol (32%), methylpavlovol (30%) and 4a-methylporiferast-22-enol (16%) as dominant sterols and with stigmasterol, ethylpavlovol and campesterol as minor sterols and Chaetoceros calcitrans contained mainly chlosterol (46%) and fucosterol (45%).
CONCLUSIONS
Sterols are an important class of membrane lipids that play vital roles in membrane stability, structure, permeability and function (Benveniste, 1986). So that, the high levels registered for the biomolecules studied sterols give to this species a special importance in for being used in food as supplements/nutraceuticals (including aquaculture). Thus, microalgae play an important role in mariculture as food for many molluscs, crustaceans and some fish. Furthermore, the best-known sterolis cholesterol, which was first discovered in human gall-stones and received the name cholesterol because of its presence in bile.
REFERENCES
- Barrett, S.M., J.K. Volkman, G.A. Dunstan and J.M. LeRoi, 1995. Sterols of 14 species of marine diatoms (Bacillariophyta). J. Phycol., 31: 360-369.
Direct Link - Fuentes, M.M., G.G. Fernandez, J.A. Perez and J.J. Guerrero, 2000. Biomass nutrient profiles of the microalga Porphyridium cruentum. Food Chem., 70: 345-353.
Direct Link - Hartmann, M.A., 1998. Plant sterols and the membrane environment. Trend Plant Sci., 3: 170-175.
Direct Link - Piironen, V., D.G. Lindsay, T.A. Miettinen, J. Toivo and A.M. Lampi, 2000. Plant sterols: Biosynthesis, biological function and their importance to human nutrition. J. Sci. Food Agric., 80: 939-966.
CrossRefDirect Link - Ponomarenko, L.P., I.V. Stonik, N.A. Aizdaicher, T.Y. Orlova, G.I. Popovskaya, G.V. Pomazkina and V.A. Stonik, 2004. Sterols of marine microalgae Pyramimonas cf. cordata (Prasinophyta), Attheya ussurensis sp. nov. (Bacillariophyta) and a spring diatom bloom from Lake Baikal. Comparative Biochem. Physiol. Part B, 138: 65-70.
Direct Link - Soudant, P., Le J. Coz, Y. Marty, J. Moal, R. Robert and J-F. Samain, 1998. Incorporation of microalgae sterols by scallop Pecten maximus (L.) Larvae. Comp. Biochem. Physiol., 119A: 451-457.
Direct Link - Volkman, J.K., 2003. Sterols in microorganisms. Applied Microbiol. Biotechnol., 60: 495-506.
Direct Link