
H. Pakniyat
Department of Crop Production and Plant Breeding, Shiraz University, Shiraz,Islamic Republic of Iran
E. Tavakol
Department of Crop Production and Plant Breeding, Shiraz University, Shiraz,Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 7 | Page No.: 1113-1117







ABSTRACT
Abstract: This research was conducted to evaluate some of the drought resistance criteria at seedling stage in wheat. A factorial experiment in a Completely Randomized Design (CRD) was used with two factors consisted of stress levels (0, -5 and -8 bar) using PEG 6000 and genotypes (10 genotypes of bread wheat; Azar 2, Gahar, Koohdasht, Bow, Zagros, Cham, Niknejad, El Neilairi, Bohoih and Giza 164) in three replications in a hydroponic condition. Stress Tolerance Index (STI), Water Use Efficiency (WUE), Biological Yield (BY), Shoot Dry Weight (SDW), Root Dry Weight (RDW), Root/Shoot weight ratio (R/S), Root Length (RL), Relative Water Content (RWC), Wilting Percentage (WP) and first and 2nd Leaves Extention Rate (LER1 and LER2) were measured at seedling stage. Increasing stress levels caused reduction in BY, SDW, RDW, RL, RWC, LER1 and LER2, but an increase in WUE, DWR, R/S and WP. Azar2, Gahar, Koohdasht, Zagros and Bow were in favorite condition in regard to STI, WUE and other criteria. Therefore, they are drought tolerant and might be suitable genotypes at water deficit conditions. Niknejad, El Neilairi and Cham were moderate and Giza 164 and Bohoih were sensitive genotypes to drought conditions. This research revealed that at -5 bar, WUE, BY, SDW, R/S and LER2 and at -8 bar, WUE, BY and WP were suitable criteria for selection of drought resistant genotypes at seedling stage.
PDF Abstract XML References Citation
How to cite this article
H. Pakniyat and E. Tavakol, 2007. Evaluation of Some Drought Resistance Criteria at Seedling Stage in Wheat (Triticum aestivum L.) Cultivars. Pakistan Journal of Biological Sciences, 10: 1113-1117.
DOI: 10.3923/pjbs.2007.1113.1117
URL: https://scialert.net/abstract/?doi=pjbs.2007.1113.1117
DOI: 10.3923/pjbs.2007.1113.1117
URL: https://scialert.net/abstract/?doi=pjbs.2007.1113.1117
INTRODUCTION
Insufficient water is a primary limitation to wheat production world-wide and cultivating drought tolerant cultivars is hampered by the lack of effective criteria (Bajji et al., 2001).
Crops are affected by drought in their lifetime but some stages such as germination, seedling stage and flowering are critical (Pessarakli, 1999). Seedling stage is an efficient stage for screening the resistant genotypes for most resistance criteria.
Knowing about root characters is very important to understand drought tolerant mechanisms in all plants (Binger and Hongwen, 2000). Root weight is effected by genotype and stress level (Shen, 1996). More growth of root can balance osmotic potential and increase water absorption ability from other layers of soil (Richards et al., 2002). Under water limited conditions, yield is considered as a function of the Water Used (WU) by the crops, the efficiency by which the water is used to produce dry matter (WUE) and the proportion of total dry matter that ends up in the grain (HI). Since it is usually assumed that there are no interactions between these factors, then an improvement in any one of them should result in an increase in yield (Passioura, 1986). This trait has genetic diversity within inter and intra species (Ehdaie and Waines, 1993). RWC is offered as an index that measured easily and rapidly to screen drought resistant genotypes (Schonfeld et al., 1988; Ritchie et al., 1990). Matin et al. (1989) measured RWC and in this way could screen resistant and sensitive genotypes at seedling stage successfully. It confirmed by measuring yield that is very time consuming criterion (Matin et al., 1989).
To study the drought stress effects on crops, synthetic macromolecules were used (Singh, 2002). Polyethylene Glycols (PEG) are neutral polymers available in a wide range of mol wt and highly soluble in water. Many investigators have used them as an osmotic agent either for whole plants or for plant tissues, cells and organelles (Plaut and Federman, 1985).
The objectives of this study were to evaluate the ability of several drought resistance criteria under different water deficit levels for identifying resistant genotypes at seedling stage in wheat.
MATERIALS AND METHODS
Plant materials: Ten bread wheat (2n = 6x = 42) genotypes collected from research centers in Fars and Yasooj Provinces in Iran (Table 1) with differed known reaction to drought stress, were tested for some important criteria at seedling stage in greenhouse of College of Agriculture, Shiraz University, Shiraz, Iran.
Hydroponic system: The seeds were sterilized by 10% sodium hypochlorite solution for 15 min and were transferred to germinator set in dark at 25°C and 60% humidity for 48 h, followed by a 8, 10 and 12 h photoperiod for 3 days to synchronize germination. Uniform-size seedlings transplanted to hydroculture pots containing half-strength Hoagland’s nutrient solution and aeriated at 15 min h-1. Experiment was arranged according to a randomized complete design with 3 replications. Plants were allowed to adjust in the hydroponics system for 2 days and then subjected to water stress using polyethylene glycol (PEG 6000-Merck) at 2 levels (ψs = -5 and -8 bar) as moderate and sever stress levels, respecively and one control treatment (0 bar). Drought stress levels were chosen based on a preliminary experiment using 6 levels (ψs= -3, -5, -6.5, -8, -9.5 and -11 bar) of water stress. To prevent osmotic shock to the plants, PEG was added gradually during a 3 days period. The concentrations of PEG for each water stress level were determined using the equation adapted from Michel and Kaufman (1973). The final ψs for each drought treatment was remained for 14 days. The volume of solution in the pots was maintained with the addition of distilled water and was recorded for calculation of WUE.
Measurements and statistical analysis: Leaf Extention Rate (LER) for the first and the second leaf (LER1 and LER2) was determined by the ratio of ΔL/Δt where ΔL = the difference between final and initial leaf length and Δt = number of days that the leaf takes to get its final length (Hay and Walker, 1994).
Two weeks after the last stage of adding PEG, 5-6 leaves of each experimental unit (pot) were separated and Relative Water Content (RWC) was estimated using the following formula:
RWC (%) = [(FW-DW)/(TW-DW)] x100
Where, FW = fresh leaf weight, TW = leaf weight after rehydration for 24 h at room temprature in the dark and DW = leaf weight after drying at 72° C for 48 h (Barrs and Weatherley, 1962).
| Table 1: | List of wheat cultivars and their drought tolerance response |
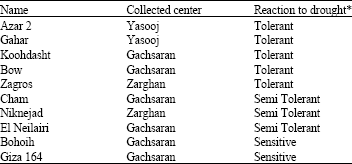 | |
| *Reaction to drought is reported by research centers which seeds were provided | |
In addition, Biological Yield (BY), Shoot Dry Weight (SDW), Root Dry Weight (RDW), Root/Shoot weight ratio (R/S), Root Length (RL) and wilting percentage were measured. WUE (Stanhil, 1986; Ehdaie and Waines, 1993; Richards et al., 2002) was determined by the proportion of total biological yield (mg) consisted of harvested shoot and root from each pot to total water used (mL) for each pot during the experiment period. The data then converted for each plant grown in pots. Stress tolerance index (STI) calculated by following formula (Fernandez, 1992).
Where Yp = biological yield of a genotype in non-stressed environment, Ys = biological yield of a genotype in stressed environment and ![]() = mean biological yield in non-stressed environment.
= mean biological yield in non-stressed environment.
Data were analyzed using SAS (1996) software. Correlation analyses was performed using Minitab (v. 13.0) statistical analysis software. Mean comparisons were performed using Duncan’s Multiple Range Test (DMRT) (Steel and Torrie, 1980). Cultivars were clustered according to their response to drought stress indices using, Farthest Neighbour Method (Kachigan, 1982).
RESULTS AND DISCUSSION
There were significant differences (p<0.01) for BY, SDW, R/S, RL, RWC and WP in different stress levels. Changes in stress levels hadn’t any significant effect on LER1. It might be due to leaf extention occurrence before starting the stress (Table 2). All the evaluated criteria showed significant differences among genotypes at all stress levels except RWC and RDW at -5 bar and non-stress treatments. Significant (p<0.01) genotype x drought stress interactions were observed for all the evaluated criteria.
Drought stress caused significant effects on wheat root system (RDW and RL). A significant reduction in RDW and RL was resulted by increasing drought stress levels (Table 2). This agrees with the results of Binger and Hongwen (2000) who found that apparently drought stress causes a reduction in root weight and length in 0-20 cm soil depth. Root/Shoot weight ratio was increased significantly by increasing drought stress level (Table 2). Marcum (1998) reported increasing in Root/Shoot weight ratio in drought stress conditions in cereals. This is an efficient way to adjust plants to drought conditions. Increasing Root/Shoot weight ratio, causes reduction in transpiration surface and root system absorb more water from wider volume of soil (Gupta, 1996).
RWC was significantly reduced by increasing drought stress levels. This agrees with the findings of Haung et al. (1998). No sigificant differences for RWC were observed between genotypes at 0 and -5 bar stress levels (Table 3). Matin et al. (1989) observed the same results for barley resistant and non-resistant genotypes. As expected, increasing drought stress levels caused a significant reduction in BY, SDW and an increase in WP (Table 2).
No measurement of WUE is reported in hydroculture system. Stress conditions (-5 and -8 bar) caused a significant increase in WUE and the difference between the two stress levels was not significant (Table 2). This is in agreement with the results obtained by Condon et al. (2002), who reported higher WUE under stress conditions compared with well-watered conditions in wheat genotypes. Crops react conservatively in facing drought conditions, hence, produce more assimilates by unit water used. In this experiment, Azar 2, Koohdasht, Bow and Zagros showed the highest WUE; Gahar, Cham, Niknejad and El Neilairi were categorized in the next rank and Bohoih and Giza 164 had the least amount at -8 bar stress (Table 4). Previous studies confirmed the effect of WUE on increasing yield (Condon et al., 2002).
Based on the correlated critertia with STI, at -8 bar stress, the cultivars were classified into 3 groups using Farthest Neighbour Method (Kachigan, 1982). These groups are: 1)-Azar2, Gahar, Koohdasht, Bow and Zagros, 2)-Niknejad, Cham and El Neilairi and 3)-Bihoih and Giza 164. In -8 bar stress level the first group showed the highest STI and WUE and the least WP, the third group showed the least amount of these criteria and the second group was in middle condition. Based on these criteria it seemes that the first group is consisted of drought tolerant, the second one semi-tolerant and the third one non-tolerant genotypes.
At -5 bar stress level STI was positively correlated with BY, SDW, WUE, RL and LER2 and negatively with R/S. STI was positively correlated with BY, SDW and WUE and negatively (p<0.05) with R/S ratio and WP at -8 bar stress level (Table 5). R/S had a significant and negative correlation with STI and BY, therefore by decreasing R/S, STI and BY will increase. WUE had significant (p<0.05) and positive correlation with BY and SDW at -5 bar stress level. Sinclair and Muchow (2001) also showed direct correlation between WUE and BY. In none of stress levels, there was significant correlation between LER and RWC. Studies had shown that, in some instances, if some parts of root is affected by drought condition, distribution of leaves are being decreased without any difference in their water condition.
| Table 2: | Average of evaluated criteria in different stress levels |
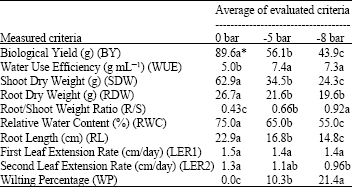 | |
| *Means followed by the same letter(s) in each row are not significantly different (DMRT, p<0.05) | |
| Table 3: | Average of evaluated criteria at -5 bar in different wheat genotypes |
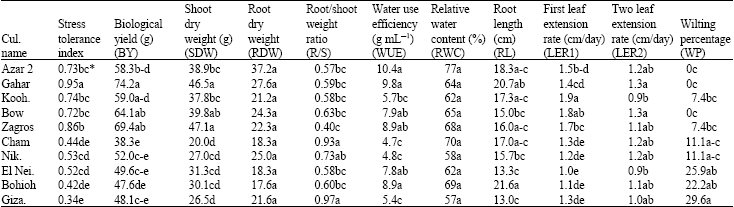 | |
| *Means followed by the same letter(s) in each column are not significantly different (DMRT, p<0.05) | |
| Table 4: | Average of evaluated criteria at -8 bar in different wheat genotypes |
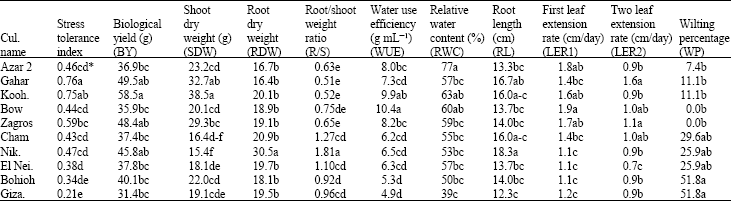 | |
| *Means followed by the same letter(s) in each column are not significantly different (DMRT, p<0.05) | |
| Table 5: | Correlation between physiological and morphological traits in seedlings of 10 genotypes of wheat. The first number corresponds to the -5 bar and the second to the -8 bar stress levels |
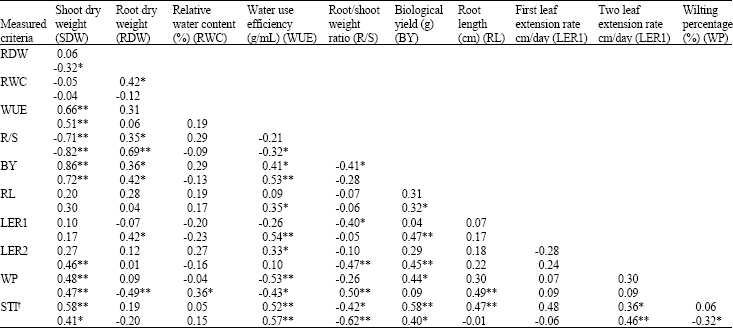 | |
| *and **show significant differences at 5 and 1% probability levels, respectively.†, Stress tolerance index | |
It’s shown that this non-hydraulically effects on leaf growth is mediated by a chemical message that originates from root and transferred to shoot by woody vessels (Blackman and Davies, 1985). We did not observe any significant relation between LER and WUE. This is in agreement with Sinclair and Muchow (2001) results who reported that change in this character causes variable responses in yield.
It can be concluded that WUE, BY, SDW, R/S ratio and LER2 at -5 bar and WUE, BY and WP at -8 bar, along with STI, are suitable criteria for screening drought tolerant genotypes at seedling stage. This conculsion seems logical based on their correlation with STI and their ability to separate genotypes according to drought stress resistance. Previous studies confirmed WUE (Ehdaie and Waines, 1993; Condon et al., 2002), WP (Nayeem and Nerkar, 1988) and R/S ratio as reliable criteria for this goal.
REFERENCES
- Bajji, M., S. Lutts and J.M. Kient, 2001. Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (Triticum durum Desf.) cultivars performing differently in arid conditions. Plant Sci., 160: 669-681.
Direct Link - Barrs, H.D. and P.E. Weatherley, 1962. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci., 15: 413-428.
Direct Link - Binger, H. and G. Hongwen, 2000. Root physiological characteristics associated with drought resistance in Tall fescue cultivars. Crop Sci., 40: 196-203.
Direct Link - Condon, A.G., R.A. Richards, G.J. Rebetzke and G.D. Farquhar, 2002. Improving intrinsic water use efficiency and crop yield. Crop Sci., 42: 122-131.
CrossRefPubMedDirect Link - Ehdaie, B. and J.G. Waines, 1993. Variation in water-use efficiency and its components in wheat: I. Well-watered pot experiment. Crop Sci., 33: 294-299.
CrossRefDirect Link - Fernandez, G.C.J., 1992. Effective selection criteria for assessing plant stress tolerance. Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, August 13-16, 1992, Shanhua, Taiwan, pp: 257-270.
Direct Link - Haung, B., J.D. Fry and B. Wang, 1998. Water relations and canopy characteristic of Tall fescue cultivars during and after drought stress. Hortic. Sci., 33: 837-840.
Direct Link - Marcum, K.B., 1998. Cell membrane thermostability and whole-plant heat tolerance of Kentucky bluegrass. Crop Sci., 38: 1214-1218.
Direct Link - Matin, M.A., J.H. Brown and H. Ferguson, 1989. Leaf water potential, relative water content and diffusive resistance as screening techniques for drought resistance in Barley. Agron. J., 81: 100-105.
CrossRefDirect Link - Michel, B.E. and R.K. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefDirect Link - Nayeem, K.A. and Y.S. Nerkar, 1988. Association of drought and heat tolerance parameters in wheat. Indian. J. Genet. Plant Breed., 48: 371-376.
Direct Link - Plaut, Z. and E. Federman, 1985. A simple procedure to overcome polyethelene glycol toxicity on whole plants. Plant Physiol., 79: 559-561.
CrossRefDirect Link - Richards, R.A., G.J. Rebetzke, A.G. Condon and A.F. van Herwaarden, 2002. Breeding opportunities for increasing the efficiency of water use and crop yield in temperate cereals. Crop Sci., 42: 111-121.
CrossRefDirect Link - Ritchie, S.W., H.T. Nguyen and A.S. Holaday, 1990. Leaf water content and gas-exchange parameters of two wheat genotypes differing in drought resistance. Crop Sci., 30: 105-111.
CrossRefDirect Link - Schonfeld, M.A., R.C. Johnson, B.F. Carver and D.W. Mornhinweg, 1988. Water relations in winter wheat as drought resistance indicators. Crop Sci., 28: 526-531.
CrossRefDirect Link - Shen, L., 1996. A comparative study on drought resistance of Lathyrus sylvesttris L. Grassland China, 3: 53-56.
PubMed - Sinclair, T.R. and R.C. Muchow, 2001. System analysis of plant traits to increase grain yield on limited water supplies. Agron. J., 93: 263-270.
Direct Link