
M. Nobakht
Department of Anatomy and Histology, Iran University, P.O. Box 14155-6183, Tehran, Iran
S. Majidzadeh
Department of Anatomy and Histology, Iran University, P.O. Box 14155-6183, Tehran, Iran
M. Fattahi
Department of Anatomy and Histology, Iran University, P.O. Box 14155-6183, Tehran, Iran
M. Samadi
Department of Anatomy and Histology, Iran University, P.O. Box 14155-6183, Tehran, Iran
P. Tabatabaeei
Department of Anatomy and Histology, Iran University, P.O. Box 14155-6183, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 7 | Page No.: 1078-1082







ABSTRACT
Abstract: The expression of neuronal nitric oxide synthase during the development of rat optic vesicle from embryonic day E14 to E18 was analyzed by histochemical procedures. The samples were frozen and cut on a cryostat and then studied by using the light microscope. Expression of nNOS was first seen on E14 in cells of Cajal-Retzius located in the marginal zone of optic vesicle. NADPH-d persisted in this layer throughout the embryonic period and began to decrease on E20. At E16, the optic vesicle has four NADPH-d positive layers. At E18, NADPH-d reactivity observed at low magnification showed five clearly defined layers. In the late stages, the most notable feature was a decrease in histochemical reaction of the marginal zone and at these stages, the layer IV showed less staining than the rest of the cortical plate. The observations suggest that nitric oxide is synthesized during embryonic life processes and this is related to maturational processes.
PDF Abstract XML References Citation
How to cite this article
M. Nobakht, S. Majidzadeh, M. Fattahi, M. Samadi and P. Tabatabaeei, 2007. Expression of Neuronal Nitric Oxide Synthase During Embryonic Development of the Rat Optic Vesicle. Pakistan Journal of Biological Sciences, 10: 1078-1082.
DOI: 10.3923/pjbs.2007.1078.1082
URL: https://scialert.net/abstract/?doi=pjbs.2007.1078.1082
DOI: 10.3923/pjbs.2007.1078.1082
URL: https://scialert.net/abstract/?doi=pjbs.2007.1078.1082
INTRODUCTION
The demonstration in 1987 of the formation of Nitric Oxide (NO) by an enzyme in the vascular endothelial cells opened up what can now be considered a new area of the biological research (Bredt, 1996; Prast and Philippu, 2001; Wang et al., 1994). Nitric oxide is an important molecule because of its role as a universal modulator of neuronal function at the synaptic level, mainly in the central nervous system. In the mammalian organism, NO is synthesized in several types of cells, such as neurons,
INTRODUCTION
The demonstration in 1987 of the formation of Nitric Oxide (NO) by an enzyme in the vascular endothelial cells opened up what can now be considered a new area of the biological research (Bredt, 1996; Prast and Philippu, 2001; Wang et al., 1994). Nitric oxide is an important molecule because of its role as a universal modulator of neuronal function at the synaptic level, mainly in the central nervous system. In the mammalian organism, NO is synthesized in several types of cells, such as neurons, endothelial cells and macrophages by a family of three isoenzymes termed nitric oxide synthases or NOS (Lopez-Costa et al., 1997). nNOS is present in neurons. It is constitutively expressed and its activity is regulated by Ca2+. Another constitutive and Ca2+-dependent type of NOS is present in endothelial cells (eNOS) of the vasculature.
Nitric oxide synthase can produce nitric oxide with the aid of NADPH in response to changes in intracellular free calcium by deimidating arginine to citrulline. The selective coexistence of citrulline-like immunoreactivity in NADPH-diaphorase positive neurons is thus consistent with NADPH diaphorase being a nitric oxide synthase (Geurges, 1999). Thus the extensive literature on the histochemistry of NADPH diaphorase should be reexamined in light of the fact that this simple histochemical technique allows the cellular localization of nitric oxide synthase in the nervous system (Dawson et al., 1991; Bruce, 1991).
In 1961, Thomas and Pearse first described the presence of large, isolated neurons containing outstanding high NADPH-diaphorase activity which were scattered throughout the cerebral cortex and basal ganglia (Schober et al., 1994).
The aim of the present study was to clarify if the rat optic vesicle cells using the NADPH-d as an indicator to the NOS-histochemistry (Rangarajan et al., 1999). Effect of NOS expression on optic vesicle development is very important on assessment abnormalities.
MATERIALS AND METHODS
In this study, 11 young adult female Spargue-Dawley rats were used. The rats were mated and the gestational age was estimated according to the day of vaginal plaque formation, to mark the first day of life. Fetuses aged 8-18 days of gestation were obtained, decapitated and washed in solution consisting of 0.1 M Phosphate Buffer Saline (PBS) in pH = 7.4. The heads were transformed into fixative for 24 h; The fixative contained fresh solution of 4% paraformaldehyde in 0.1 M PBS, pH = 7.4, at 4deg; C. Subsequently the heads were kept in solution consisting of 15% sucrose in PBS for 1 h and then in 30% sucrose at 4deg; C for 2 h. They were then frozen and transverse and longitudinal sections to 10 μ M were cut on a cryostat. Sections were collected on gelatin-coated slides. They were later rinsed in 0.1 M PBS (pH = 8.0) for 10 min. NADPH-diaphorase histochemistry was performed by incubating the sections in a solution consisting of 30 μ M malic acid, 1 μ M manganese hydrochloride, 0.2 μ M nitro blue tetrazolium chloride, 1 μ M NADPH, 0.5% Triton X-100 in buffer of Tris-HCl in pH 7.4, at 37deg; C, for 90 min in a dark place. Sections were mounted in entellan and photographs were taken.
RESULTS
At E13, the diencephalic vesicle is composed of only two layers: the ventricular zone, where all neurons are generated and the marginal zone, where the first-generated neurons are setting (Fig. 1).
NADPH-d histochemistry was not observed in the diencephalic vesicle at this stage in either transverse or longitudinal sections.
At E14, the diencephalic vesicle is greatly enlarged and increased in thickness, mainly due to growth of the ventricular zone (Fig. 2). The marginal zone is also thicker. An incipient intermediate zone between the ventricular zone and marginal zone is observed. The whole marginal zone of the diencephalic vesicle was populated by NOS-positive cells. There were two chief types of NADPH-d histochemistry cells (Fig. 3), large cells in the middle part of marginal zone with long tangentially oriented processes, which showed the morphological characteristics of Cajal-Retzius cells and externally situated round cells with processes oriented upward.
At E16, the diencephalic vesicle has four layers: the ventricular zone, which occupies roughly one-third of the thickness, the intermediate zone, with easily visible fibers coursing along it, marginal zone and the cortical plate and more prominent in the lateral than in the dorsal aspect of the diencephalic vesicle.
At E18, NADPH-d reactivity is observed at low magnification showed five clearly defined layers (Fig. 4).
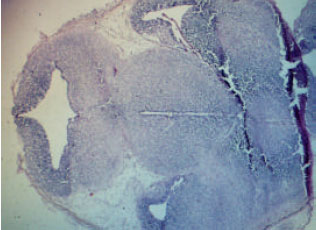 | |
| Fig. 1: | Frontal section through diencephalen. Rat embryo at E13, H and E staining |
The marginal zone is densely labeled and the underlying cortical plate showed a band of stained cells in which nearly every cell was stained, so that it was difficult to identify single cells.
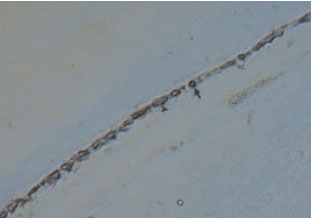
In the late stage, the most notable feature was a decrease in the histochemical reaction of the marginal zone. Horizontally coursing NADPH-d histochemical activity fibers could also be observed in the marginal zone. Also NADPH-d reactivity was observed in endothelial cells lining the larger blood vessels, but it was not present in endothelial cells of sinusoidal capillaries or the small vessels (Fig. 5).
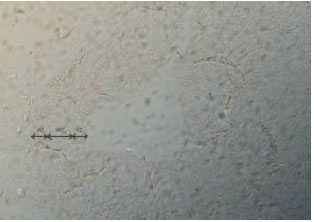 | |
| Fig. 2: | Frontal section through the optic rat embryo at E14 optic vesicle Viewed at higher magnification. At this stage, the diencephalic vesicle continues to grow and its thickness increases. The marginal zone has been also thicker and because of growth of the ventricular zone, the intermediate zone is more prominent. Cryostat section |
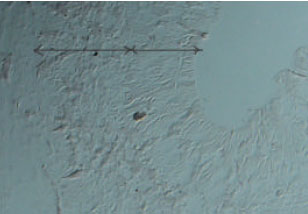 | |
| Fig. 3: | Frontal section showing the diencephalic vesicle of a rat embryo at E14 at high magnification. The picture shows two layers of marginal zone and ventricular zone. There are NOS-positive round cells in the marginal zone. The nerve fibers are also stained. At the ventricular zone, only nerve fibers are weakly stained. Cryostat section |
 | |
| Fig. 4: | Photomicrograph showing the marginal zone rat embryo at E18Staining is observed in a group of highly active cells. There are two kinds of NOS-positive cells: big chief cells in the middle part of the marginal zone with long processes having the characteristics of Cajal-Retzius cells and small round cells in the lateral portion. Cryostat section |
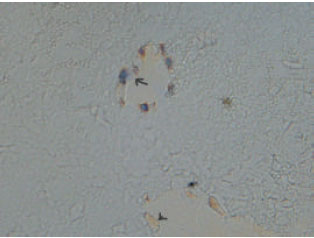 | |
| Fig. 5: | NADPH-d histochemistry in the rat cells embryo at E19. Here are also shown the cells without reaction. Cryostat section |
DISCUSSION
The first embryonic neurons to express nNOS in the diencephalic vesicle were those situated in the marginal zone. According to the studies of Bayer and Altman (1990) using autoradiography, tritiated thymidine, the Cajal-Retzius cells that are characteristic of this layer are generated at E13 and E14. Cajal-Retzius cells play a significant role throughout the development of the cerebral cortex. According to Marva-Padilla, every cell generated during the development of cerebral cortex established contact with Cajal-Retzius cells of the marginal zone. As maturation proceeds, only pyramidal neurons retain and expand their original connections with the marginal zone, while other neuronal types lose them. Ogawa et al. (1995) have shown that Cajal-Retzius cells express the protein reelin.
Only a few authors have studied nNOS or NADPH-d expression in Cajal-Retzius cells during embryonic development (Bayer and Altman, 1985; Bredt and Synder, 1994; Yan et al., 1996). These cells are seldom seen in mature animals. In the adult rat, layer I cells are not stained for nNOS (Brown, 2001), but others have recently shown nNOS-immunopositive neurons of Cajal-Retzius morphology can be found in layer I of aged rats (Uttenthal in press). These results suggest that the lack of nNOS expression by these cells of layer I is a prolonged, but transient feature of adult life (Florenzano and Guglielmitti, 2000).
In the present study, the intense histochemistry reaction and morphological characteristics of these cells persisted during the embryonic period and only began to decrease at E20, when neuronal migration is coming to an end. We observed that cells populating the marginal zone were abundant from E20 to the first postnatal stages.
Autoradiographic studies have shown that these cells are generated between E12 and E14; we have observed this phenomenon at E14 in the external part of the marginal zone. We suggest that NO may be involved in directing the ingrowing axons and in the migrational process.
During the development of the diencephalic vesicle, numerous afferent fibers reach the cortex from subcortical structures (Bayer and Altman, 1985; Bayer and Altman, 1991). These do not express choline acetyl transferase until the second postnatal week, where as NGF (Nerve Growth Factor) is expressed in the embryonic basal forebrain as early as E13 (Koh and Loy 1989; Weitzberg and Lundberg, 1998; Santacana et al., 1998).
Another important feature at E17 is the presence of migrating cells expressing nNOS in the intermediate zone. The expression of nNOS or NADPH-d and the role of NO cell migration are controversial issues. Some authors (Schilling et al., 1994) suggest that, in the cerebellum, cells express nNOS only when migration is completed. Studies on slice cultures of rat cerebellum (Tanakas et al., 1994; Riccio et al., 2006) have shown that granule cell migration is inhibited by N-nitro-L-arginin (L-NNA), indicating that NO was involved in cell migration and in the differentiation of granule cells.
In this study, the presence of migrating cells expressing nNOS in the intermediate zone of the diencephalic vesicle suggests that NO is involved in migration process.
From E18, a new embryonic layer is clearly observed in Nissl-stained sections, the supra ventricular zone. At the main source of neurogenesis, the ventricular zone, shrinks, many cells are reduced in the supra ventricular zone. This layer does not completely disappear after birth but continues to generate cells. It has been thought that the germinal cells in the supra ventricular zone produce only astrocytes and oligodentrocytes (Moncada et al., 1991). At E18, nNOS-reactive fibers were observed in the intermediate zone (Hanel and Hensey, 2006).
The present study de scribes the expression of nNOS in the diencephalic vesicle during embryonic development. Neuronal NOS expression may be considered to be correlated with the production of NO and may thus provide indirect evidence for a role of NO in development. During the embryonic stages, the role of NO is probably different from that in the adult. This is suggested by much greater expression of nNOS found in the cerebral cortex during the embryonic stages than in later life (Wang et al., 1994).
First of all, in the diencephalic vesicle, the neuronal elements expressing nNOS are much more abundant in embryonic stages than in neonates or adults (Bredt and Suyder, 1994; Giuili et al., 1994; Northington et al., 1996; Drever et al., 2004). In the adult nervous system, the role of NO in physiological conditions is the productoin a neurotransmitter or neuromodulator (Dawson and Dawson, 1996; Koliatsos, 2004). However, the large amount of nNOS in the embryonic diencephalic vesicle suggests an important role at that stage.
The results presented suggest that NO expression whenever there is an activity related to maturational processes.
ACKNOWLEDGMENTS
We would like to express our heartful thanks to Mr. Negahdar for his valuable guidance. Furthermore, we are grateful to Ms Hoseini for type processing and Mr. Gheibi and Mr. Zohrehvand for providing us with special facilities.
endothelial cells and macrophages by a family of three isoenzymes termed nitric oxide synthases or NOS (Lopez-Costa et al., 1997). nNOS is present in neurons. It is constitutively expressed and its activity is regulated by Ca2+. Another constitutive and Ca2+-dependent type of NOS is present in endothelial cells (eNOS) of the vasculature.Nitric oxide synthase can produce nitric oxide with the aid of NADPH in response to changes in intracellular free calcium by deimidating arginine to citrulline. The selective coexistence of citrulline-like immunoreactivity in NADPH-diaphorase positive neurons is thus consistent with NADPH diaphorase being a nitric oxide synthase (Geurges, 1999). Thus the extensive literature on the histochemistry of NADPH diaphorase should be reexamined in light of the fact that this simple histochemical technique allows the cellular localization of nitric oxide synthase in the nervous system (Dawson et al., 1991; Bruce, 1991).
In 1961, Thomas and Pearse first described the presence of large, isolated neurons containing outstanding high NADPH-diaphorase activity which were scattered throughout the cerebral cortex and basal ganglia (Schober et al., 1994).
The aim of the present study was to clarify if the rat optic vesicle cells using the NADPH-d as an indicator to the NOS-histochemistry (Rangarajan et al., 1999). Effect of NOS expression on optic vesicle development is very important on assessment abnormalities.
MATERIALS AND METHODS
In this study, 11 young adult female Spargue-Dawley rats were used. The rats were mated and the gestational age was estimated according to the day of vaginal plaque formation, to mark the first day of life. Fetuses aged 8-18 days of gestation were obtained, decapitated and washed in solution consisting of 0.1 M Phosphate Buffer Saline (PBS) in pH = 7.4. The heads were transformed into fixative for 24 h; The fixative contained fresh solution of 4% paraformaldehyde in 0.1 M PBS, pH = 7.4, at 4C. Subsequently the heads were kept in solution consisting of 15% sucrose in PBS for 1 h and then in 30% sucrose at 4C for 2 h. They were then frozen and transverse and longitudinal sections to 10 μM were cut on a cryostat. Sections were collected on gelatin-coated slides. They were later rinsed in 0.1 M PBS (pH = 8.0) for 10 min. NADPH-diaphorase histochemistry was performed by incubating the sections in a solution consisting of 30 μM
malic acid, 1 μM manganese hydrochloride, 0.2 μM nitro blue tetrazolium chloride, 1 μM NADPH, 0.5% Triton X-100 in buffer of Tris-HCl in pH 7.4, at 37C, for 90 min in a dark place. Sections were mounted in entellan and photographs were taken.
RESULTS
At E13, the diencephalic vesicle is composed of only two layers: the ventricular zone, where all neurons are generated and the marginal zone, where the first-generated neurons are setting (Fig. 1).
NADPH-d histochemistry was not observed in the diencephalic vesicle at this stage in either transverse or longitudinal sections.
At E14, the diencephalic vesicle is greatly enlarged and increased in thickness, mainly due to growth of the ventricular zone (Fig. 2). The marginal zone is also thicker. An incipient intermediate zone between the ventricular zone and marginal zone is observed. The whole marginal zone of the diencephalic vesicle was populated by NOS-positive cells. There were two chief types of NADPH-d histochemistry cells (Fig. 3), large cells in the middle part of marginal zone with long tangentially oriented processes, which showed the morphological characteristics of Cajal-Retzius cells and externally situated round cells with processes oriented upward.
At E16, the diencephalic vesicle has four layers: the ventricular zone, which occupies roughly one-third of the thickness, the intermediate zone, with easily visible fibers coursing along it, marginal zone and the cortical plate and more prominent in the lateral than in the dorsal aspect of the diencephalic vesicle.
At E18, NADPH-d reactivity is observed at low magnification showed five clearly defined layers (Fig. 4).
| |
| Fig. 1: | Frontal section through diencephalen. Rat embryo at E13, H and E staining |
The marginal zone is densely labeled and the underlying cortical plate showed a band of stained cells in which nearly every cell was stained, so that it was difficult to identify single cells.
In the late stage, the most notable feature was a decrease in the histochemical reaction of the marginal zone. Horizontally coursing NADPH-d histochemical activity fibers could also be observed in the marginal zone. Also NADPH-d reactivity was observed in
|
| Fig. 2: | Frontal section through the optic rat embryo at E14 optic vesicle Viewed at higher magnification. At this stage, the diencephalic vesicle continues to grow and its thickness increases. The marginal zone has been also thicker and because of growth of the ventricular zone, the intermediate zone is more prominent. Cryostat section |
|
| Fig. 3: | Frontal section showing the diencephalic vesicle of a rat embryo at E14 at high magnification. The picture shows two layers of marginal zone and ventricular zone. There are NOS-positive round cells in the marginal zone. The nerve fibers are also stained. At the ventricular zone, only nerve fibers are weakly stained. Cryostat section |
|
| Fig. 4: | Photomicrograph showing the marginal zone rat embryo at E18Staining is observed in a group of highly active cells. There are two kinds of NOS-positive cells: big chief cells in the middle part of the marginal zone with long processes having the characteristics of Cajal-Retzius cells and small round cells in the lateral portion. Cryostat section |
| |
| Fig. 5: | NADPH-d histochemistry in the rat cells embryo at E19. Here are also shown the cells without reaction. Cryostat section |
endothelial cells lining the larger blood vessels, but it was not present in endothelial cells of sinusoidal capillaries or the small vessels (Fig. 5).
DISCUSSION
The first embryonic neurons to express nNOS in the diencephalic vesicle were those situated in the marginal zone. According to the studies of Bayer and Altman (1990) using autoradiography, tritiated thymidine, the Cajal-Retzius cells that are characteristic of this layer are generated at E13 and E14. Cajal-Retzius cells play a significant role throughout the development of the cerebral cortex. According to Marva-Padilla, every cell generated during the development of cerebral cortex established contact with Cajal-Retzius cells of the marginal zone. As maturation proceeds, only pyramidal neurons retain and expand their original connections with the marginal zone, while other neuronal types lose them. Ogawa et al. (1995) have shown that Cajal-Retzius cells express the protein reelin.
Only a few authors have studied nNOS or NADPH-d expression in Cajal-Retzius cells during embryonic development (Bayer and Altman, 1985; Bredt and Synder, 1994; Yan et al., 1996). These cells are seldom seen in mature animals. In the adult rat, layer I cells are not stained for nNOS (Brown, 2001), but others have recently shown nNOS-immunopositive neurons of Cajal-Retzius morphology can be found in layer I of aged rats (Uttenthal in press). These results suggest that the lack of nNOS expression by these cells of layer I is a prolonged, but transient feature of adult life (Florenzano and Guglielmitti, 2000).
In the present study, the intense histochemistry reaction and morphological characteristics of these cells persisted during the embryonic period and only began to decrease at E20, when neuronal migration is coming to an end. We observed that cells populating the marginal zone were abundant from E20 to the first postnatal stages.
Autoradiographic studies have shown that these cells are generated between E12 and E14; we have observed this phenomenon at E14 in the external part of the marginal zone. We suggest that NO may be involved in directing the ingrowing axons and in the migrational process.
During the development of the diencephalic vesicle, numerous afferent fibers reach the cortex from subcortical structures (Bayer and Altman, 1985; Bayer and Altman, 1991). These do not express choline acetyl transferase until the second postnatal week, where as NGF (Nerve Growth Factor) is expressed in the embryonic basal forebrain as early as E13 (Koh and Loy 1989; Weitzberg and Lundberg, 1998; Santacana et al., 1998).
Another important feature at E17 is the presence of migrating cells expressing nNOS in the intermediate zone. The expression of nNOS or NADPH-d and the role of NO cell migration are controversial issues. Some authors (Schilling et al., 1994) suggest that, in the cerebellum, cells express nNOS only when migration is completed. Studies on slice cultures of rat cerebellum (Tanakas et al., 1994; Riccio et al., 2006) have shown that granule cell migration is inhibited by N-nitro-L-arginin (L-NNA), indicating that NO was involved in cell migration and in the differentiation of granule cells.
In this study, the presence of migrating cells expressing nNOS in the intermediate zone of the diencephalic vesicle suggests that NO is involved in migration process.
From E18, a new embryonic layer is clearly observed in Nissl-stained sections, the supra ventricular zone. At the main source of neurogenesis, the ventricular zone, shrinks, many cells are reduced in the supra ventricular zone. This layer does not completely disappear after birth but continues to generate cells. It has been thought that the germinal cells in the supra ventricular zone produce only astrocytes and oligodentrocytes (Moncada et al., 1991). At E18, nNOS-reactive fibers were observed in the intermediate zone (Hanel and Hensey, 2006).
The present study de scribes the expression of nNOS in the diencephalic vesicle during embryonic development. Neuronal NOS expression may be considered to be correlated with the production of NO and may thus provide indirect evidence for a role of NO in development. During the embryonic stages, the role of NO is probably different from that in the adult. This is suggested by much greater expression of nNOS found in the cerebral cortex during the embryonic stages than in later life (Wang et al., 1994).
First of all, in the diencephalic vesicle, the neuronal elements expressing nNOS are much more abundant in embryonic stages than in neonates or adults (Bredt and Suyder, 1994; Giuili et al., 1994; Northington et al., 1996; Drever et al., 2004). In the adult nervous system, the role of NO in physiological conditions is the productoin a neurotransmitter or neuromodulator (Dawson and Dawson, 1996; Koliatsos, 2004). However, the large amount of nNOS in the embryonic diencephalic vesicle suggests an important role at that stage.
The results presented suggest that NO expression whenever there is an activity related to maturational processes.
ACKNOWLEDGMENTS
We would like to express our heartful thanks to Mr. Negahdar for his valuable guidance. Furthermore, we are grateful to Ms Hoseini for type processing and Mr. Gheibi and Mr. Zohrehvand for providing us with special facilities.
REFERENCES
- Bredt, D.S. and S.H. Snyder, 1994. Nitric oxide synthase neurons in embryonic cerebral cortical plate, sensory ganglia and olfactory epithelium. Neuron, 13: 301-313.
Direct Link - Bruce, T., 1991. Neuronal NADPH diaphorase is a nitric oxide synthase. Proc. Natl. Acad. Sci. USA., 88: 2811-2814.
Direct Link - Dawson, E.D., T.M. Bredt, D.S. Fotuhi, P.M. Hwang and S.H. Synder, 1991. Nitric oxide synthase and neuronal NADPH-diaphorase are identical in brain and peripheral tissues. Proc. Natl. Acad. Sci. USA., 88: 7797-7801.
Direct Link - Drever, J., M. Schleicher, A. Tappe, K. Schilling and T. Kuner et al., 2004. Nitric oxide synthase NOS-interacting protein interacts with neuronal NOS and regulates its distribution and activity. J. Neurosci., 24: 10454-10465.
Direct Link - Florenzano, F. and V. Guglielmotti, 2000. Selective nicotinamide adenine dinucleotide phosphate-diaphorase histochemical labeling of Muller radial processes and photoreceptors in the earliest stages of retinal development in the tadpole. Neurosci., Lett., 292: 187-190.
Direct Link - Geurges, P., 1999. Apoptosis during development of the human retina: Relationship to foveal development and retinal synaptogenesis. J. Compar. Neurol., 113: 198-208.
Direct Link - Giuili, G., A. Luzi, M. Poyard and G. Guellaen, 1994. Expression of mouse brain soluble guanylyl cyclase and NO synthase during ontogeny. Dev. Brain Res., 81: 269-283.
Direct Link - Hanel, M.L. and C. Hensey, 2006. Eye and neural defects associated with loss of GDF6. BMC Dev. Biol., 6: 43-43.
Direct Link - Koliatsos, V.E., T.M. Dawson, A. Kecojevic, Y. Zhou, Y.F. Wang and K.X. Huang, 2004. Cortical interneurons become activated by differentiation and instruction the apoptosis of pyramidal neurons. Proc. Natl. Acad. Sci. USA., 101: 14264-14269.
Direct Link - Lopez-Costa, J.J., J. Goldstein and J.P. Saavedra, 1997. Neuronal and macrophagic NOS isoforms distribution in normal rat retina. Neurosci. Lett., 232: 155-158.
Direct Link - Moncada, S., R.M. Palmer and E.A. Higgs, 1991. NO: Physiology, pathophysiology and pharmacology. Pharmacol. Rev., 43: 109-142.
Direct Link - Northington, F.J., R.C. Koehler, R.J. Traystman and L.J. Martin, 1996. Nitric oxide synthase 1 and nitric oxide 3 protein expressions is regionally and temporally regulated in fetal brain. Dev. Brain Res., 95: 1-14.
Direct Link - Ogawa, M., K. Miyata, Nakajima, K. Yagyu and M. Seike et al., 1995. The reeler gene-associated antigen on Cajal-Retzius neurons is a crucial molecule for laminar organization of corticalneurons. Neuron, 14: 899-912.
Direct Link - Prast, H. and A. Philippu, 2001. Nitric oxide as modulator of neuronal function. Prog. Neurobiol., 64: 51-68.
Direct Link - Rangarajan, R., Q. Gong and U. Gaul, 1999. Migration and function of glia in the developing Drosophila eye. Development, 126: 3285-3292.
Direct Link - Riccio, A., R.S. Alvania, B.E. Lonze, N. Ramanan and T. Kim et al., 2006. A nitric oxide signaling pathway controls CREB-mediated gene expression in neurons. Mol. Cell., 21: 283-294.
Direct Link - Santacana, M., L.O. Uttenthal, M.L. Bentura, A.P. Fernander and J. Serrano, 1998. Expression of neuronal nitric oxide synthase during embryonic development of the rat cerebral cortex. Dev. Brain Res., 111: 205-222.
Direct Link - Schilling, K., H.H. Schmidt and S.L. Baader, 1994. Nitric oxide synthase expression reveals compartments of cerebellar granule cells and suggests a role for mossy fibers in their development. Neuroscience, 59: 893-903.
Direct Link - Schober, A., C.R. Malz, W. Schober and D.L. Meyer 1994. NADPH-diaphorase in the central nervous system of the larval Lamprey (Lampetra planeri). J. Comp. Neurol., 345: 94-104.
Direct Link - Tanakas, M., S. Yoshida, M. Yano and F. Hanaoka, 1994. Roles of endogenous nitric oxide in cerebellar cortical development in slice cultures. Neuroport, 5: 2049-2052.
Direct Link - Uttenthal, L.O., D. Alonso, A.P. Fernandez, R.O. Campbell and M.A. Moro et al., 1998. Neuronal and inducible nitric oxide synthases and nitrotyrosine immunoreactivities in the cerebral cortex of aging rats. Microsc. Res Tech., 43: 75-88.
Direct Link - Wang, Z.Z., L.J. Stensaas, D.S. Bredt, B. Dinger and S.J. Fidone, 1994. Localization and actions of nitric oxide in the cat carotid body. Neuroscience, 60: 275-286.
Direct Link - Weitzberg, E. and J.O. Lundberg, 1998. Nonenzymatic nitric oxide production in humans. Nitric Oxide, 2: 1-7.
Direct Link - Yan, X.X., L.J. Garey and L.S. Jen, 1996. Prenatal development of NADPH-diaphorase-reactive neurons in human frontal cortex. Cereb. Cortex, 6: 737-745.
Direct Link