
A.M. Kheirallah
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
T.I. Tantawi
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
A.H. Aly
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
Z.A. El-Moaty
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 7 | Page No.: 1001-1010







ABSTRACT
Rearing of L. sericata and C. albiceps in pure cultures of a variety of densities, at four constant temperature regimes, demonstrated an inverse relationship between egg density and the total development (F2,6 = 264.35 and F2,6 = 71.87 For L. sericata and C. albiceps, respectively p<0.05), survivorship (F2,9 = 68.41; p< 0.001 for L. sericata and F2,9 = 12.35; p< 0.05 for C. albiceps) and adult size (For L. sericata: males F2,9 = 58.94 and females F2, 9 = 140.09. For C. albiceps: males F2, 9 = 43.62 and females F2, 9 = 167.99. In all cases p<0.001) of both species. Addition of C. albiceps reduced the development time and survivorship of L. sericata at a variety of densities and proportions at 17 and 23°C. While at 29 and 35°C, a complete elimination of L. sericata was recorded. Neither egg density nor temperature has a significant effect on the sex ratio of the resulting adults. It seems that the interaction between these species is a highly asymmetric case of competition.
PDF Abstract XML References Citation
How to cite this article
A.M. Kheirallah, T.I. Tantawi, A.H. Aly and Z.A. El-Moaty, 2007. Competitive Interaction Between Larvae of Lucilia sericata (Meigen) and Chrysomya albiceps (Wiedemann) (Diptera: Calliphoridae). Pakistan Journal of Biological Sciences, 10: 1001-1010.
DOI: 10.3923/pjbs.2007.1001.1010
URL: https://scialert.net/abstract/?doi=pjbs.2007.1001.1010
DOI: 10.3923/pjbs.2007.1001.1010
URL: https://scialert.net/abstract/?doi=pjbs.2007.1001.1010
INTRODUCTION
Competition in nature is difficult to detect and defining of competition is not straightforward (Paul, 1993). The natural history of carrion flies is thought to include intense competition for larval food (Suenaga, 1959a, b; Beaver, 1973, 1977; Cornaby, 1974; Baxter and Morrison, 1983; So and Dugeon, 1990; Wells and Greenberg, 1992a, b). Almost every aspect of their breeding biology suggests strong selection for rapid location and consumption of this patchy and empheral resource (Beaver, 1984; Hanski, 1987).
Intra- and inter-specific larval competition for food result in larval mortality, reduction in size and weight of individual larvae and pupae and undersized adults with less fecundity and short life (Ullyett, 1950; Putman, 1977; Williams and Richardson, 1983; Shahein, 1986; Goodbrod and Goff, 1990; Omar, 1992; Reis et al., 1994; Von Zuben, et al., 2001).
The degree to which blowflies species suffer loss of population from the effects of intra and interspecific competition on a carcass is determined by their inherent growth characteristics and by the degree to which they are adapted to withstand the adverse conditions engendered by overcrowding of the larval populations on the available food supply (Ullyett, 1950).
Intra- and inter-specific competition in two species of calliphorids; the common green bottle blow fly Lucilia sericata and the African hairy maggot blow fly Chrysomya albiceps can be easily cultured and studied in detail under laboratory conditions.
The blow fly L. sericata is a communicative eusynathrope, which is widespread throughout the major zoogeographical regions, but is not yet cosmopolitan (Greenberg and Povolny, 1971; Smith, 1986; Spradbery, 1991). It is a Holarctic species distributed over the warmer regions of the temperate zone (Hall and Wall, 1995) and now the dominant species in urban and suburban districts of Australia and Africa (Zumpt, 1965; Wall et al., 1992). L. sericata is a facultative ectoparasite, which acts as the primary agent of sheep myiasis in Britain and parts of continental Europe (Smith, 1931; Davies, 1934; MacLeod, 1943; Wall et al., 1992; Hall and Wall, 1995; Smith and Wall, 1997a), in South Africa (Hepburn, 1943) and Australia (Norris, 1959).
Chrysomya albiceps is a hemisynathropic species, which prefers high temperatures and humidity (Greenberg and Povolny, 1971). This tropical and subtropical species is very common and abundant in Africa, southern Europe, tropical and subtropical Arabia, India and recently central and South America (Zumpt, 1965; Guimarães et al., 1978, 1979; Mariluis, 1980, 1981, 1983; Leite et al., 1983; Baumgartner and Greenberg, 1984; Baumagartner, 1988; Spradbery, 1991; Hall and Smith, 1993; Grassberger et al., 2003).
In field carrion studies, in Egypt, Tantawi et al. (1996) found that L. sericata was able to breed successfully in carrion in fall, winter and spring. While in summer, little or no breeding in carrion occurred when the adult population was most abundant. This behavior displayed by L. sericata was attributed as to avoid intense predation of its larvae by third instars of C. albiceps in summer, during which larval development of both species proceeds synchronously. Therefore, it was found necessary to explore this aspect by a series of laboratory experiments.
MATERIALS AND METHODS
Maintenance of laboratory fly colonies: Larvae and adults of the two species were collected from exposed rabbit carcasses at Moharrem Bey District, Alexandria, Egypt. Larvae and adults were identified to species after Tantawi and El-Kady (1997). The flies were held in fly screen cage (50x50x50 cm) at 25±2°C. Adults were supplied with water, granulated sucrose and powdered milk.
Experimental procedures: Gravid females of L. sericata or C. albiceps were allowed to oviposit on minced meat placed in black 35-mm film cups. This provided a dark and moist environment preferred by female flies for oviposition. Eggs were collected within 30 min of oviposition, so that they would be the same age.
Clumps of eggs were separated by a gentle shaking for 10 min in a tube of 0.1 M NaOH (Sandeman et al., 1987). Unseperated eggs, which float unlike the separated eggs and the solution were decanted. The remaining eggs were rinsed twice by filling the tube with distilled water, which then was decanted when the eggs had settled. The eggs were poured with a small amount of water onto a paper towel marked with a grid and counted under a dissecting microscope (Model 41 THRU 48). A piece of towel with the desired number of eggs was then cut out and placed with the eggs down on the meat of the experimental jar. Care was taken that the towel did not dry before the eggs were placed in each experimental jar.
Containers used in these experiments were 1 L jars. Each jar filled, in the following order, with about 5 cm height of moistened sawdust, 140 g of minced meat, the desired combination of calliphorid eggs and dry sawdust to within 2 cm of the top. The mouth of the jar was then covered with a fine mesh cloth, sealed by a rubber band and placed into an incubator at one of the four desired temperature regimes (17, 23, 29 or 35±0.5°C). The quantity of meat used in these experiments was selected according to previous experiments of competition conducted by Ullyett (1950) and Wells and Greenberg (1992a). To estimate hatching success, a number of leftover eggs (at least in hundreds) were kept moist in petri dishes next to the jars. In every case, the hatching success percentage should exceed 90%.
Intra-specific competition (pure cultures): Three treatments of 40, 200 and 1000 eggs of each species were selected. The quantities of food available per larva for these treatments were 3.5, 0.7 and 0.14 g, respectively.
Inter-specific competition (mixed cultures): Nine combinations of 20, 100 or 500 eggs of L. sericata or C. albiceps were selected and performed in this manner:
L20 C20, L20 C100, L20 C500, L100 C20, L100 C100, L100 C 500, L500 C20, L500 C100 and L500 C500.
Development under different temperature regimes: In this study, the minimum duration of total development (from egg to adult eclosion) of each species at each of the four studied temperature regimes in pure and mixed cultures was recorded. As the larvae grew the mean temperature within the center of actively feeding maggots was recorded at regular intervals throughout the entire feeding period by inserting the probe of a calibrated mercury thermometer in the core of the maggot mass. The time range of the adult eclosion was determined from periodical observations at 6 h intervals. This procedure was in triplicates for each egg density at each of the tested temperature regimes.
Survivorship criteria of adults: The effect of larval competitionon the resulting adults was studied as follows: adult flies were killed by refrigerating after 12 h of observing the first fly to eclose, sorted by species and counted. Sexes were determined by examining the flies under a stereomicroscope (Model OLY 31) (Smith, 1986). Samples of adult flies of each sex were randomly collected, dried at 50°C and weighed.
Statistical analysis: Statistical analysis was performed using the program SAS (Institute Inc (1988), Cary, NC, USA), data were analyzed statistically by using Two-ways analysis of variance (ANOVA) and a t-test to detect differences between means (Sokal and Rohlf, 1981).
RESULTS
Development rate: For pure cultures, Table 1 data reveled significant effects (p<0.05) of temperature and egg density on the development of both species (For L. sericata: F3,6 = 38427.31 and F2,6 = 264.35, respectively. For C. albiceps: F3,6 = 20736.35 and F2,6 = 71.87, respectively).
At all tested temperatures, the maggot mass temperature of each species showed a regular pattern of increase as the number of eggs was increased. In cultures of lower egg densities (40 eggs), the differences between the ambient and maggot mass temperature were slight and showed no detectable pattern. On the other hand, in cultures of higher densities (200 and 1000), strong peaks in temperature were observed. A maximal temperature of 11°C for L. sericata and 13.5°C for C. albiceps above ambient was observed in the culture with density of 1000 eggs.
For mixed cultures, the results showed that at higher temperatures (29, 35°C) complete elimination of L. sericata was observed from all egg combinations, except at a ratio of 25L: 1C (L500:C20), where L. sericata was able to complete development and survive. Analysis of data found a significant decrease in the development time of L. sericata with increasing intra- and inter-specific competition in the cultures with 20, 100 and 500 of C. albiceps (In all cases p<0.05). Concerning the development time of C. albiceps, results of ANOVA showed a significant decrease in the development time with increasing only intraspecific competition (In all cases p<0.05). On the other hand, when the egg density of C. albiceps held constant and that of L. sericata increased, insignificant effects of increasing interspecific competition was indicated (In all cases p>0.05).
Analysis of the data also revealed that at lower temperatures (17, 23°C) the total development time of L. sericata in pure cultures was slower than in mixed cultures of corresponding total egg densities, in all cases p<0.05. On the other hand, at 17 and 23°C, the total development time of C. albiceps was lengthened significantly (p<0.05) in mixed cultures (L100C100, L500C500) than in pure cultures of egg densities (200, 1000). Also at both 29 and 35°C, a significant increase in the development of C. albiceps was observed in pure cultures of egg density (1000) than in mixed cultures of egg combinations (L500C500), in all cases p<0.05 (Table 1).
Survivorship: The proportion surviving was arcsine transformed and subjected to ANOVA. For pure cultures, the results revealed that adult survivorship of both species declined significantly as the initial egg number was increased (F2,9 = 68.41; p<0.001 for L. sericata and F2,9 = 12.35; p<0.05 for C. albiceps). It is clear that there was a consistent pattern of higher survivorship at 29°C for both species (Table 2).
For mixed cultures, no statistically true effect of intra- and inter-specific competition on the survivorship of L. sericata at cultures of 20 and 100 (In all cases p>0.05) but significantly decreased in culture with 500 eggs (p<0.05). On the other hand, there was a significant decrease in the survivorship of C. albiceps with increasing only the intraspecific competition (In all cases p<0.05) (Table 2).
| Table 1: | Minimal total development times (in hours) of Lucilia sericata and Chrysomya albiceps raised in pure and mixed cultures of a variety of densities and proportions at different temperature regimes |
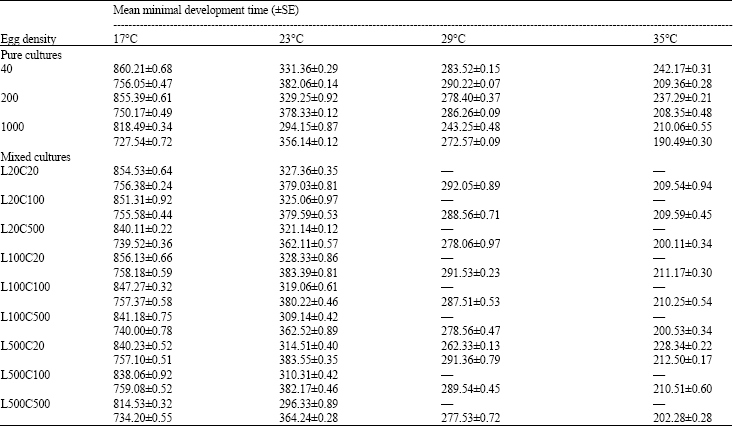 | |
| Upper figures for Lucilia sericata, Lower figures for Chrysomya albiceps. Data obtained from 3 replicate rearings | |
| Table 2: | Percentage of adult survivorship of Lucilia sericata and Chrysomya albiceps raised in pure and mixed cultures of a variety of densities and proportions at different temperature regimes |
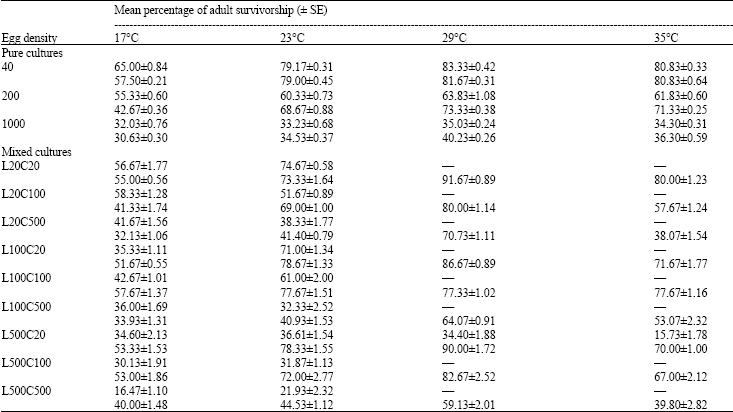 | |
| Upper figures for Lucilia sericata, Lower figures for Chrysomya albiceps, Data obtained from 3 replicate rearings | |
| Table 3: | Percentage of female adults of Lucilia sericata and Chrysomya albiceps raised in pure and mixed cultures of a variety of densities and proportions at different temperature regimes |
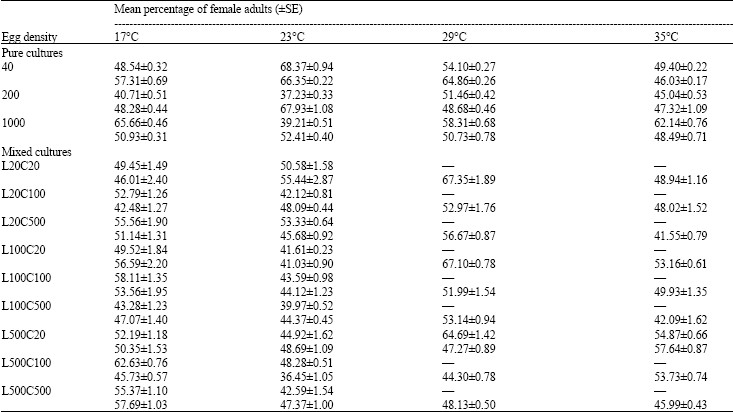 | |
| Upper figures for Lucilia sericata, Lower figures for Chrysomya albiceps, Data obtained from 3 replicate | |
Analysis of the data revealed a significant increase in mortality of L. sericata, at 17 and 23°C, in the mixed cultures than in pure cultures of identical total egg densities, in all cases p<0.05. On the other hand, at higher temperatures (29, 35°C), the survivorship of C. albiceps was significantly greater in the mixed culture than that in the pure cultures, in all cases p<0.05 (Table 2).
| Table 4: | Adult size (dry weight in mg) of Lucilia sericata and Chrysomya albiceps raised in pure and mixed cultures of a variety of densities and proportions at different temperature regimes |
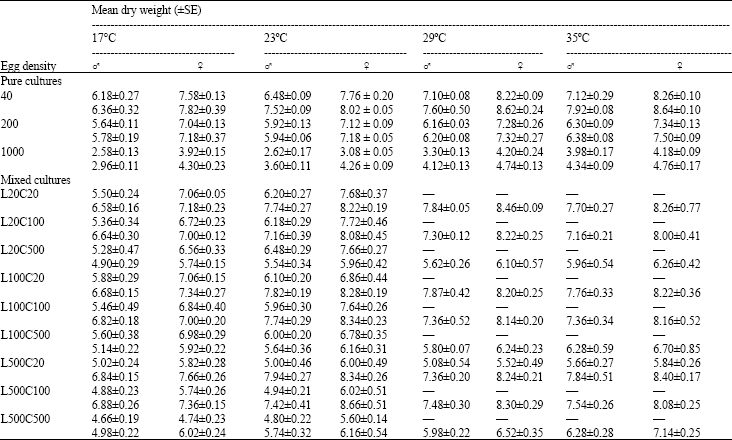 | |
| Upper figures for Lucilia sericata, Lower figures for Chrysomya albiceps, Data obtained from 3 replicate rearings | |
Sex ratio: For both pure and mixed cultures, it can be noticed that the percentage of surviving females of both species was nearly around the value of 50%. Analysis of data revealed that the percentage of surviving females were insignificantly affected by either egg density or temperature and there were insignificant differences in the percentages females of each species from pure or mixed cultures, in all cases p>0.05 (Table 3).
Size: In pure cultures, there was a significant decrease in adult size of both sexes of each species as the initial number of eggs was increased (For L. sericata: males F2,9 = 58.94 and females F2, 9 = 140.09. For C. albiceps: males F2, 9 = 43.62 and females F2, 9 = 167.99. In all cases p<0.001) (Table 4).
For mixed cultures, the adult size of L. sericata significantly decreased with increasing both intra- and inter-specific competition (In all cases p<0.05). For C. albiceps, the adult size significantly decreased only with increasing intraspecific competition. In contrast, a significant increase in the adult size of C. albiceps was recorded with increasing interspecific competition in cultures of 500 eggs (Table 4).
DISCUSSION
Development rate: Due to the rapid deterioration of the carrion habitat, the relative success of larvae will depend on how quickly they attain the minimum weight for viable pupation (Ullyett, 1950; Levot et al., 1979). For both species, the development time decreased with increasing the number of eggs was increased. Reduction of the development time as the initial number of eggs was increased has been observed in other dipterans (Ullyett, 1950; Baxter et al., 1973; Baxter and Morrison, 1983; Hutton and Wasti, 1980; So and Dudgeon, 1989a, b and 1990; Goodbrod and Goff, 1990; Ribeiro et al., 1993; Saunders and Bee, 1995).
It has been shown in the study by Baxter and Morrison (1983) that fly larvae reared in crowded conditions may benefit from the presence of others through elevation of temperature, accumulation of digestive enzymes and physical alterations of the carcasses which will facilitate feeding. The present study reported excess heat in maggot mass of L. sericata and C. albiceps which peaked at 11.0 and 13.5°C, respectively above the ambient temperature. In the study of flies development on dead bodies, a process of larvae mass self heating was observed owing to metabolic heat release and results in an up to 50% shortening of the development in a certain period of pre-imago stages, compared to the time on the environmental temperature data (Marchenko, 1973; Zvereva and Marchenko, 1987). Elevation of maggot mass temperature is common in many carrion flies (Marchenko 1985; Lord et al., 1986; Early and Goff, 1986; Goodbrod and Goff, 1990; Cianci and Sheldon 1990). The present study adds support to the view of Goodbrod and Goff (1990) who mentioned that maggot mass could be another factor for consideration when estimating postmortem intervals (PMI), based on maggot development.
The current results of mixed cultures indicate that, at a variety of densities and proportions at temperatures 29 and 35°C, L. sericata was completely eliminated. This result is in accordance to Hutton and Wasti (1980) and Furman et al. (1959). This elimination could be explained in view of Tantawi et al. (1996) that L. sericata breed successfully in carrion in fall, winter and spring. However, in summer, no breeding in carrion occurred when the adult population of L. sericata was most abundant. This behaviour of L. sericata avoided intense predation of its larvae by third instar of C. albiceps, in summer, during which larval development of both species proceeds synchronously. Larval predation by C. albiceps has been reported, in laboratory experiments, by several authors (Coe, 1978; Gagné, 1981; Erzinçlioglu and Whitcombe, 1983; Faria and Godoy, 2001).
It also found that the development of C. albiceps was not significantly influenced in cultures where C. albiceps density was held constant and that of L. sericata increased. This result contradicts that obtained by Wells and Greenberg (1992a) who recorded a prolongation in the development of C. rufifacies with increasing interspecific density of C. macellaria. Comparison between pure and mixed cultures indicates that, at all the density level tested, the development of L. sericata was shortened significantly in mixed than in pure cultures, the reverse was true for C. albiceps, at higher egg density. We believe that prolongation of C. albiceps development in mixed cultures may be attributed to the ability of C. albiceps, under food scarceness, to change its behaviour attacking L. sericata larvae. Thus the feeding larvae of C. albiceps spent much longer time during the feeding process. This result is in agreement with Wells and Greenberg (1992a) and Aguiar-Coelho and Milward-de-Azevedo (1998)
Survivorship: Presented data of single-species cultures indicated that survivorship at each tested temperature decreased as a function of increasing egg densities in both species. This result in accordance with (Ullyett, 1950; Miller, 1964; Klomp, 1964; Wasti et al., 1975and b; Moon, 1980; Shahein, 1986; So and Dudgeon, 1989a; Ribeiro, 1990; Goodbrod and Goff, 1990; Godoy et al., 1993; Von Zuben et al., 1993; Smith and Wall, 1997b; Aguiar-Coelho and Milward-de-Azevedo, 1998; Silva et al., 2003). In contrast, Wells and Greenberg (1992a) reported insignificant effect of density on survival of C. macellaria and C. rufifacies in pure culture experiments. Explanations for decreasing survival at higher egg densities have been forward by De Jong, (1976); Baxter and Morrison (1983); Hanski (1987); Lominicki (1988); Godoy et al. (1993) and Reis et al. (1994, 1996 and 1999). These authors have emphasized the importance of increasing levels of exploitative competition for limited resources among larvae.
In double-species cultures, the high survivorship of C. albiceps and the zero survivorship of L. sericata, in all egg combinations at both temperatures 29 and 35°C are indicative of the predatory behaviour of C. albiceps third instar against L. sericata larvae. The greater survival ability of C. albiceps larvae was attributed to their greater competitive ability and predatory activity in mixed cultures against larvae from other dipteran (Ullyett, 1950 and Marckenko, 1985). At temperatures 17 and 23°C, for mixed cultures, as the proportion of a species is decreased the influence of intraspecific interaction on survivorship becomes less important and that of interspecific interaction more important. Wells and Greenberg (1992a) found that the interaction between C. rufifacies and C. macellaria larvae significantly decreased C. macellaria survivorship but no effect in the reverse direction. Goodbrod and Goff (1990) and Reis et al. (1999) mixed C. rufifacies and C. megacephala, C. putoria and C. macellaria, respectively. They found that the interaction increased the survivorship of the former species and decreased that of the latter species.
Sex ratio: In this study, neither egg density nor temperature has a significant effect on the sex ratio of both species raised in pure and mixed cultures. Similar observation was recorded for C. rufifacies and C. macellaria (Wells and Greenberg, 1992a). It is evident from previous studies, that the density levels did not show any remarkable effect on the sex ratio of various dipteran species; M. domestica (Sullivan and Sokal, 1963);
L. sericata (Klomp, 1964); D. melanogaster (Miller, 1964); M. autumnalis (Moon, 1980); C. erythrocephala (Shahein, 1986). Amano (1989) attributed this due to the density would affect male and female larvae to the same extent as for pupariation. In contrast, in a similar study, Omar (1992) observed that the sex ratio of C. albiceps was significantly affected by egg density since the males number predominates with the appearance of a few females as the larval densities increase.
Size: The present results indicated that adult size of both species reduced in higher densities than in lower densities. Declines in adult size in favour of larval survivorship is common in many Diptera colonizing ephemeral habitats (such as carrion, dung and decaying fruit; Ullyett, 1950; Kitaoka, 1957; Bakker, 1961; Sullivan and Sokal, 1963; Manoukas and Tsiropoulos, 1977; Atkinson, 1979; Butlin and Day, 1984; Sigurjonsdottir, 1984). Explanation for smaller body size, mediated by competition has already been demonstrated by Sullivan and Sokal (1963). So and Dudgeon (1989b) reported that reduced food individual ration to Boettcherisca formosensis larvae in crowded cultures produced undersized adults.
Comparison between adult size in pure and mixed cultures shows that, for L. sericata, the size of both sexes decreased more rapidly with increasing egg density in the mixed cultures than in the pure cultures. The reverse was true for C. albiceps. Hence, in terms of a reduction in adult size, the data suggest that L. sericata experienced higher levels of competition in the mixed than in the pure cultures.
It was shown here that L. sericata is competitively inferior species, although coexistence between L. sericata and C. albiceps does occur in nature. Tantawi et al. (1996), reported that L. sericata was able to breed successfully in carrion in fall, winter and spring, but during summer months, no breeding in carrion occurred. This behaviour of L. sericata avoided intense predation by C. albiceps third instar. Therefore, extinction of L. sericata is unlikely in field due to avoiding breeding in summer during which both species developed synchronously.
REFERENCES
- Aguiar-Coelho, V.M. and E.M.V. Milward-de-Azevedo, 1998. Combined rearing of Cochliomyia macellaria (Fabr.), Chrysomya megacephala (Fabr.) and Chrysomya albiceps (Wied.) (Diptera: Calliphoridae) under laboratory conditions. J. Applied Entomol., 122: 551-554.
Direct Link - Atkinson, W.D., 1979. A field investigation of larval competition in domestic Drosophila. J. Anim. Ecol., 48: 91-102.
Direct Link - Baumgartner, D.L. and B. Greenberg, 1984. The genus Chrysomya (Diptera: Calliphoridae) in the new world. J. Med. Entomol., 21: 105-113.
Direct Link - Cianci, T.J. and J.K. Sheldon, 1990. Endothermic generation by blow fly larvae Phormia regina developing in pig carcasses. Bull. Soc. Vector Ecol., 15: 33-40.
Direct Link - Cornaby, B.W., 1974. Carrion reduction by animals in contrasting tropical habitats. Biotropica, 6: 51-63.
CrossRefDirect Link - Early, M. and M.L. Goff, 1986. Arthropod succession patterns in exposed carrion on the island of O'Ahu, Hawaiian Islands, USA. J. Med. Entomol., 23: 520-523.
CrossRefDirect Link - Faria, L.D.B. and W.A.C. Godoy, 2001. Prey choice by facultative predator larvae of Chrysomya albiceps (Diptera: Calliphoridae). Mem. Ins. Oswaldo Cruz., 96: 875-878.
Direct Link - Godoy, W.A.C, S.F. Dos-Reis, C.J. Von-Zuben and O.B. Ribeiro, 1993. Population dynamics of Chrysomya putoria (Wied.) (Dipt., Calliphoridae). J. Applied Entomol., 116: 163-169.
Direct Link - Goodbrod, J.R. and M.L. Goff, 1990. Effects of larval population density on rates of development and interactions between two species of Chrysomya (Diptera: Calliphoridae) in laboratory culture. J. Med. Entomol., 27: 338-343.
CrossRefPubMedDirect Link - Grassberger, M., E. Friedrich and C. Reiter, 2003. The blow fly Chrysomya albiceps (Wiedemann) (Diptera: Calliphoridae) as a new indicator in Central Europe. Int. J. Legal Med., 117: 75-81.
Direct Link - Hall, M. and R. Wall, 1995. Myiasis of humans and domestic animals. Adv. Parasitol., 35: 257-334.
CrossRefPubMedDirect Link - Omar, A. H., 1992. Effect of larval population density on the dynamics of growth in Chrysomya albiceps (Wiedemann) I. Effect on the variations observed in subsequent development of pupae and adults. J. Egypt. Soc. Zool., 22: 521-531.
Direct Link - Reis, S.F., M.A. Teixeira, F.J. Von-Zuben, W.A.C. Godoy and C.J. Von-Zuben, 1996. Theoretical dynamics of experimental populations of introduced and native blow flies (Dipt., Calliphoridae). J. Med. Entomol., 33: 537-544.
Direct Link - Reis, S.F., C.J. von Zuben and W.A.C. Godoy, 1999. Larval aggregation for food in experimental population of Chrysomya putoria (Wied.) and Cochliomyia macellaria (F.) (Diptera: Calliphoridae). J. Applied Entomol., 123: 485-489.
Direct Link - Sandeman, R.M., B.J. Collins and P.R. Carnegie, 1987. A scanning electron microscope study of L. cuprina larvae and the development of blow fly strike in sheep. Int. J. Parasit., 33: 759-765.
Direct Link - Saunders, D.S. and A. Bee, 1995. Effects of larval crowding on size and fecundity of the blow fly Calliphora vicina (Diptera: Calliphoridae). Eur. J. Entomol., 92: 615-622.
Direct Link - Silva, I.C.R., P.F.A.Mancera and W.A.C. Godoy, 2003. Population dynamics of Lucilia eximia (Diptera: Calliphoridae). J. Applied Entomol., 127: 2-6.
Direct Link - Smith, K.E. and R. Wall, 1997. The use of carrion as breeding sites by the blow fly Lucilia sericata and other Calliphoridae. Med. Vet. Entomol., 11: 38-44.
Direct Link - Smith, K.E. and R. Wall, 1997. A symmetric competition between larvae of the blow flies Calliphora vicina and Lucilia sericata in carrion. Ecol. Entomol., 22: 468-474.
Direct Link - So, P.M. and D. Dudgeon, 1990. Interspecific competition among larvae of Hemipyrellia ligurries (Calliphoridae) and Boettcherisca formosensis (Sarcophagidae) (Diptera). Res. Popul. Ecol., 32: 337-348.
Direct Link - Tantawi, T.I., E.M. El-Kady, B. Greenberg and H.A. El-Ghaffar, 1996. Arthropod succession on exposed rabbit carrion in Alexandria, Egypt. J. Med. Entomol., 33: 566-580.
Direct Link - Tantawi, T.I. and E.M. El-Kady, 1997. Identification of third instar larvae of forensically important flies (Diptera: Calliphoridae, Sarcophagidae and Muscidae) in Alexandria, Egypt. J. Egypt. Ger. Zool., 23: 1-20.
Direct Link - Von Zuben, C.J., S.F. Dos-Reis, J.B. R. Val, W.A.C. Godoy and O.B. Ribeiro, 1993. Dynamics of a mathematical model of Chrysomya megacephala (Diptera: Calliphoridae). J. Med. Entomol., 30: 443-448.
Direct Link - Von-Zuben, C.J., F.J. Von-Zuben and W.A.C. Godoy, 2001. Larval competition for patchy resources in Chrysomya megacephala (Diptera: Calliphoridae): Implications of the spatial distribution of immatures. J. Applied Entomol., 125: 537-541.
Direct Link - Wall, R., N.P. French and K.L. Morgan, 1992. Blow fly species composition in sheep myiasis in Britain. Med. Vet. Entomol., 6: 177-178.
Direct Link - Wells, J.D. and B. Greenberg, 1992. Laboratory interaction between introduced Chrysomya rufifacies and native Cochliomyia macellaria (Diptera: Calliphoridae). Environ. Entomol., 21: 640-645.
CrossRefDirect Link - Wells, J.D. and B. Greenberg, 1992. Interaction between Chrysomya rufifacies and Cochliomyia macellaria (Diptera: Calliphoridae): The possible consequences of an invasion. Bull. Entomol. Res., 82: 133-137.
Direct Link - Zumpt, F., 1965. Myiasis in Man and Animals in the Old World: A textbook for Physicians, Veterinarians and Zoologists. Butterworths Publishing Company, London, Pages: 267.
CrossRefDirect Link