
Yusuf Bozkurt
Department of Aquaculture, Faculty of Fisheries, Mustafa Kemal University,31040 Hatay, Turkey
Selcuk Secer
Department of Fisheries and Aquaculture, Faculty of Agriculture, Ankara University, 06110 Ankara, Turkey
Neslihan Bukan
Department of Biochemistry, Faculty of Medicine, Gazi University, 06510 Ankara, Turkey
Ergun Akcay
Deartment of Reproduction and Artificial Insemination, Faculty of Veterinary, Ankara University, 06110 Ankara, Turkey
Necmettin Tekin
Deartment of Reproduction and Artificial Insemination, Faculty of Veterinary, Ankara University, 06110 Ankara, Turkey
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 5 | Page No.: 940-944







ABSTRACT
This study investigated the relationships between body condition, seminal plasma composition and physical parameters of Salmo trutta fario in order to find out biomarkers for semen quality. Seminal plasma contained 79.0±15.15 mmol L-1 Na+, 46.8±9.84 mmol L-1 K+, 3.5±0.67 mg dL-1 Ca+4, 3.5±0.80 (mEqL-1 Mg++, 14.8±5.15 mgdl glucose, 3.0±9.42 g dL-1 protein, 19.2±18.57 mg dL-1 cholesterol, 5.4±3.17 mg dL-1 triglyceride and 3.0±9.42 mg dL-1 urea. Semen volume was 3.90±1.48 mL, spermatozoa motility 81.0±10.74%, duration of spermatozoa movement 97.4±15.23 s., spermatozoa density 9432.5±3762.07 x109 mL-1, total spermatozoa density 35102.4±19137.5 x109 and semen pH 7.6±0.39. There were significant positive correlations between weight and protein (r=0.752, p<0.05), weight and cholesterol (r = 0.832, p<0.01), length and protein (r = 0.729, p<0.05), length and cholesterol (r = 0.761, p<0.05), volume and cholesterol (r = 0.667, p<0.05), volume and urea (r = 0.753, p<0.05), density and total density (r = 0.704, p<0.05), total density and calcium (r = 0.676, p<0.05), Na+ and K+ (r = 0.822, p<0.01), Mg++ and protein (r = 0.932, p<0.01), protein and cholesterol (r = 0.882, p<0.01), cholesterol and urea (r = 0.885, p<0.01). Significantly negative correlation were found between K+ and pH (r = -0.891, p<0.01), Ca++ and pH (r = -695, p<0.05) and Na+ and urea (r = -0.798, p<0.01). The phenotypic correlation between body weight and length was found highly significant (r = 0.984, p<0.01).
PDF Abstract XML References Citation
How to cite this article
Yusuf Bozkurt, Selcuk Secer, Neslihan Bukan, Ergun Akcay and Necmettin Tekin, 2006. Relationship Between Body Conditon, Physiological and Biochemical Paramaters in Brown Trout (Salmo trutta fario) Sperm. Pakistan Journal of Biological Sciences, 9: 940-944.
DOI: 10.3923/pjbs.2006.940.944
URL: https://scialert.net/abstract/?doi=pjbs.2006.940.944
DOI: 10.3923/pjbs.2006.940.944
URL: https://scialert.net/abstract/?doi=pjbs.2006.940.944
INTRODUCTION
In commercial fish production the evaluation of sperm quality is of interest to increase the efficiency of artificial fertilization. The fish farming industry has been more focused on the quality of eggs or larvae rather than that of sperm, even though the quality of both gametes may affect fertilization success and larval survival. Sperm quality in farmed fish may be affected by diferent components of broodstock husbandry, during collection and storage of sperm prior to fertilization. In some species, poor sperm quality can be a limiting factor in their culture. However, even when fertilization succes is high, differences in sperm quality between males when mixed sperm from multiple males is used may severely reduce the apparent population size and may affect the future genetic integrity of the stock. In Salmonids, spermatogenesis is a seasonal event and chemical and physical properties of semen may change since all spermatozoa are eliminated by the end of the reproductive season (Lahnsteiner et al., 1993). Sperm consists of seminal plasma and spermatozoa. Seminal plasma contains substances that support sperm cells. Some substances reflect the functioning of the reproductive system and spermatozoa (Ciereszko and Dabrowski, 2000). The main role of seminal plasma is to create an optimal environment for spermatozoa storage. Information on the composition of seminal plasma and other biological fluids can be used to make media for use as a diluent or for gamete storage. Better knowledge of sperm components is important to understanding events leading to production of good quality gametes and to identifying factors that disturb semen function. Salmo trutta forms resident populations in the upper streams of rivers and occurs in North Africa, Europe, West Asia and Anatolia (Geldiay and Balik, 1988) and it is an important potential species for recreational fishery. However, in most parts of these areas, river systems have undergone great changes in their ecology and morphology in recent years and river damming and degradation of spawning habitats have caused a decline in the stocks of Salmo trutta.
Considered together, available data indicate that there are individual differences in sperm quantity and quality that must be identified to select the most valuable fish for broodstock. Up to now, differences in sperm production and quality among male Salmo trutta fario have not been evaluated. To obtain exact knowledge about biomarkers for sperm quality, the present study investigates the relationship between physical spermatological properties and seminal plasma composition.
MATERIALS AND METHODS
Broodstock care and collection of semen: Ten mature (2+ years old) male Salmo trutta fario (total weight 1.475±0.28 kg, total length 31.85±5.82 cm) were used as semen donors. The broodstock were held in 1.5 x 1 x 10 m raceways under a natural photoperiod regime and fed with a commercial trout diet (containing 40% protein) at 2% of their body weight per day. Water temperature varied between 7-8°C during spawning season. Semen from each fish was collected into 25 mL calibrated glass beakers by abdominal massage. Only pure samples uncontaminated with slime, faeces or blood were used.
Determination of spermatozoa motility, duration of movement, spermatozoa density and pH: A 10 μL sample was taken from each semen batch and placed on a microscope slide and 100 μL activation solution (0.3% NaCl) was added to determine spermatozoa motility. The percentage of motility was defined as the percentage of progressively motile spermatozoa within each activated sample. Progressively motile spermatozoa were defined as actively swimming in a forward motion. Sperm cells that vibrated in place were not considered to be motile and observations were made within thirty minutes of semen collection. Duration of spermatozoa movement was estimated using a sensitive chronometer. The spermatozoa density was calculated using the hemocytometric method. For this aim, semen samples were left on Thoma’s hemocytometer undisturbed for a few minutes prior to counting to allow sperm cells to settle. Counts were conducted at x200 magnification and expressed as x109 mL-1. Semen pH was also measured with standart pH electrodes within thirty minutes of sampling.
Determination of seminal plasma composition: Seminal plasma of the semen from each fish was collected after centrifugation of the semen at 3000 rpm for 10 min at room temperature and stored in Eppendorf vials. Na+, K+, Ca++, Mg++, glucose, protein, cholesterol, triglyceride and urea levels were determined by Abbott-Aeroset autoanalyser (USA) using original kits.
Statistical analyses: Results are presented as means±SD. Differences between parameters were analysed by one-way analyses of variance (ANOVA). Significant means were subjected to a multiple comparison test (Duncan) at α = 0.05 level. All analysis were carried using the SPSS 10 for Windows statistical software package.
RESULTS AND DISCUSSION
The spermatological properties of the semen are presented in Table 1. The composition of the seminal plasma ions and metabolites are presented in Table 2. It is interesting to note that significant positive correlations were found between weight and protein (r = 0.752, p<0.05), weight and cholesterol (r = 0.832, p<0.01), length and protein (r = 0 .729, p<0.05) and length and cholesterol (r = 0.761, p<0.05). Also, the phenotypic correlation between body weight and length was found highly significant (r = 0.984, p<0.01). Correlations between body weight-length, spermatological and biochemical properties are presented in Table 3.
Mean sperm volume of brown trout was found rather low when compared to rainbow trout reported by Geffen and Evans (2000), Akçay et al. (2002a) and Tekin et al. (2003). The differences may be due to the feeding conditions and regime, environmental factors or spawning time. Spawning time of brown trout begin rather earlier and shorter (only two months) than rainbow trout. Short spermatogenesis period may be one reason for the low semen production. In the present study, the mean spermatozoa motility agreed with the findings of Schmidt-Baulain and Holtz (1989) and Tekin et al. (2003) but not with Levanduski and Cloud (1988) on rainbow trout. In the case of movement duration; the finding was rather similar with that of Akcay et al. (2002a) but not with Munkittrick and Moccia (1987) and Levanduski and Cloud (1988). Spermatozoa motility varies in vigor and duration not only among males but also within an individual male depending on ripeness (Akcay et al., 2002b). Most studies on fish species have shown that the duration and motility of sperm may vary seasonally (Benau and Terner, 1980). The findings on spermatozoa density in the present study agrees with the finding of Akcay et al. (2002a) and Tekin et al. (2003) but differed from Munkittrick and Moccia (1987), McNiven et al. (1993), Ciereszko and Dabrowski (1993) and Secer et al. (2004).
| Table 1: | Spermatological parameters of Salmo trutta fario semen (n = 10) |
 | |
| Table 2: | Means±standard deviation (n = 3) for seminal plasma ion and metabolite composition of Salmo trutta fario semen |
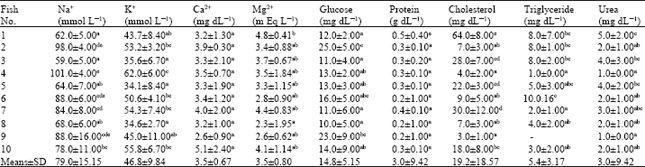 | |
| Different superscripts in a column indicate significant differences at p<0.05 | |
| Table 3: | Correlations between body weight-length, spermatological properties and seminal plasma composition of Salmo trout fario semen |
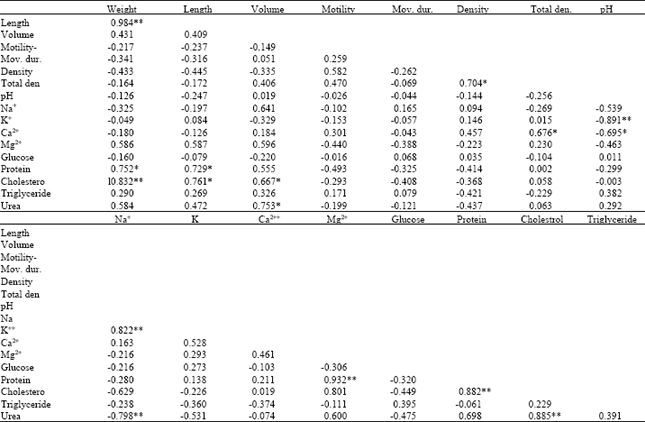 | |
| *Correlation is significant at p<0.05 **Correlation is significant at p<0.01 | |
The differences may be due to feeding conditions, age, environmental factors or dilution ratios. Sperm pH was found in the agrees with the findings of Akcay et al. (2002a) and range of generally confirmed (Piironen, 1985; Munkittrick and Moccia, 1987).
Seminal plasma of brown trout has a similar Na+ content with rainbow trout (80 mmol L; Secer et al., 2004), common carp (75 mmol L-1; Morisawa et al., 1983) but lower than perch (124 mmol L-1; Lahnsteiner et al., 1995), catfish (164 mmol L -1; Tan-Fermin et al., 1999) and muskellunge (129 mmol L-1; Lin et al., 1996). However, the K+ content was higher than reported for perch (10 mmol L-1; Lahnsteiner et al., 1 995), catfish (18 mmol L-1; Tan- Fermin et al., 1999) and muskellunge (28 mmol L-1; Lin et al., 1996) and similar with rainbow trout (46 mmol L-1; Secer et al., 2004) but lower than common carp (70 mmol L-1; Morisawa et al., 1983). Electrolytes (such as Na+ and K+) ensure the viability of sperm. The K+ ion has a role in keeping spermatozoa in the quiescent state (Baynes et al., 1981). Low levels of Na+ and K+ ions are associated with low percentages of motile spermatozoa and such semen are considered low quality. However, the high levels of motility, Na+ and K+ determined in the present study do not support this situation. In addition, it can be concluded that Ca++ and Mg++ contribute significantly to the ionic composition of seminal plasma of brown trout. White and Macleod (1963) indicated that protein has a protective role. High protein concentration (3.0±9.42 g dL-1) indicates that high protein amounts are necessary for brown trout semen. Contamination of sperm with contaminants can seriously affect the biochemical parameters of seminal plasma and if undetected may produce false results. In the present study, the low levels of urea indicate that only a little contamination is available. The presence of glucose in seminal plasma has been connected to the high energy demand of the testes during spermatogenesis or to lipid synthesis of spermatozoa (Soengas et al., 1993). Various lipid classes have been found in seminal plasma and their levels vary greatly among fish species, such as 0.007 g L-1 for Arctic charr (Piironen and Hyvarinen, 1983), 1.00 g L-1 for Euroasian perch (Piironen, 1994) and 2.55 mg dL-1 for rainbow trout (Secer et al., 2004). In addition, low levels of triglycerides were found in the seminal plasma of cyprinids (Lahnsteiner et al., 1994) and rainbow trout (Secer et al., 2004) like brown trout in the present study. Triglycerides serve as energy resources for rainbow trout spermatozoa during immotile storage and during the regeneration phase after motility (Lahnsteiner et al., 1993). Low triglycerides and glycerol levels could therefore be indicative of inadequate energy resources, reduced motility rate and fertilization capacity. According to Piironen (1994), seminal plasma lipids are associated with metabolism in spermatozoa. While cholesterol was found in the seminal plasma of freshwater fish (Billard et al., 1995), there is little information about its role. Lipids and cholesterol might have a protective effect against environmental changes (especially water temperature) when semen was released.
The phenotypic correlation between body weight and length was found very high (r = 0.984) and statistically important (p<0.01). The correlations between body weight-length and spermatological and biochemical parameters generally were found as negative and insignificant except protein and cholesterol. Similarly, Billard et al., (1974) reported an insignificant correlation (r = 0.11) between total number of spermatozoa and body weight in rainbow trout. In addition, assuming a low relationship between body weight and concentration of spermatozoa as reported in Atlantic salmon (Kazakov, 1981). Sperm production and quality can be affected by both fish size and physiological status. On the other hand, relationships found as low and insignificant in this experiment between fish size and sperm quality indices indicate that physical condition of mature fish has not a influence on sperm quality. According to results of the present study it can be concluded that sperm quality may be affected from the effects of genetics, diet and environmental stres (toxicants, water quality, fish density). In addition, according to Campbell et al. (1992), fish held in captivity experience conditions, like the present experimet, that often increase stress and lead to reduced gamete quantity.
In conclusion, the present study indicates that, mature males releasing sperm with low motility and low density should be culled from the broodstock. Reducing the number of male broodstock maintained for spawning can significantly improve hatchery efficiency and minimize feed costs. Our data can be used to select high quality mature males for fertilizing eggs in a commercial aquaculture operation. In addition to these, the information on sperm physiology obtained from the present study can lead to more efficient gamete management and increased fry yields and aid suitability of sperm for frozen or liquid preservation.
REFERENCES
- Billard, R., J. Cosson, G. Perchec and O. Linhart, 1995. Biology of sperm and artificial reproduction in carp Aquaculture, 129: 95-112.
CrossRefDirect Link - Ciereszko, A. and K. Dabrowski, 2000. In vitro effect of gossypol acetate on yellow perch (Perca flavescens) spermatozoa. Aquat. Toxicol., 49: 181-187.
Direct Link - Tekin, N., S. Secer, E. Akcay and Y. Bozkurt, 2003. Cryopreservation of rainbow trout (Oncorhynchus mykiss) semen. Israeli J. Aquacult. Bamidgeh, 55: 208-212.
Direct Link