
N. Gulsoy
Department of Biology, Faculty of Science and Art, Marmara University, G�ztepe, İstanbul, Turley
Y. Aytekin
Department of Histology and Embryology, Faculty of Istanbul Medicine, Istanbul University, �apa, Istanbul, Turkey
R. Yuce
Department of Biology, Faculty of Science and Art, Marmara University, G�ztepe, İstanbul, Turley
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 5 | Page No.: 935-939







ABSTRACT
The follicular epithelium and theca of oocytes in Onchorynchus mykiss two years old, differentiates during the initial primary growth phase. The follicular cells are squamous and the thecal cells are consisted of two layers. When the oocytes developed, follicular cells become cuboidal. These cells show characteristics typical of protein- or glycoprotein- producing cells and exhibit dilated intercellular spaces. Formation egg envelope in O. mykiss begins in the previtellogenic oocytes as a layer of amorphous electron dense materials laid down the oolemma. During the vitellogenesis, several microvilli developed between the follicular epithelium and oolemma rapidly. The morphology of egg envelope in the trout reflects its oviparous nature.
PDF Abstract XML References Citation
How to cite this article
N. Gulsoy, Y. Aytekin and R. Yuce, 2006. Changing of Follicular Epithelium During Oogenesis in Rainbow Trout
(Oncorhynchus mykiss, W.), Studied by Light and Electron Microscopy. Pakistan Journal of Biological Sciences, 9: 935-939.
DOI: 10.3923/pjbs.2006.935.939
URL: https://scialert.net/abstract/?doi=pjbs.2006.935.939
DOI: 10.3923/pjbs.2006.935.939
URL: https://scialert.net/abstract/?doi=pjbs.2006.935.939
INTRODUCTION
Egg production is annual event and controlled by the various factors. In teleost development of the oogonia transform the follicle complex. The follicle consists of oocyte and follicular epithelium. Basal lamina and two cell layers (theca interna and theca externa) surrounding the follicle is described by Grier, (2000). Reviews of oocyte growth in teleost have been given by Tyler and Sumpter (1996), Wallace and Selman (1981) and de Vlaming (1983). Oogenesis in trout, Oncorhynchus mykiss W., has studied by light microscopy by Nomura (1962), van den Hurk and Peute (1979), Tyler et al. (1990) and Krisfalusi et al. (2000). The fine structure of oocytes has been reviewed by Guraya (1986).
The present study describes the follicular complex in two years old trout, living in the pools of Sarikiz stream, was studied in Kütahya, Turkey. During the primary and secondary growth phases the changing of follicular epithelium was described by light and transmission electron microscopy.
MATERIALS AND METHODS
Research material, two years old trout, Oncorhynchus mykiss, were sampled randomly on a montly basis between July 2001 and June 2002 from an established domesticated stock with a natural spawning period of November-December were used at sarikiz stream in Kütahya, Turkey. Each month 7-10 fish were sampled for a total 103 individuals. After anesthetization with 0.035% M.S. 222, fish were measured to total length to the nearest 0.5 cm, body weight to the nearest 5 g and ovary mass 0.01 g. The range of total length of female fish used in the present study was 26.50-43.00 cm. In the calculation of gonadosomatic index (GSI) the following formula was employed.
The ovaries for light microscopy were fixed in bouin solution immediately. After dehydration tissues were embedded in either Histo sec 56-58 pF (Merck) or hydroxyethyl methacrylate and sectioned at 4-7 μm. Transverse and longitudinal sections were obtained in ovaries. The following dyes were used for samples kept in Histosec; hematoxylin-eosin as general coloring, azan trichromics for collogen, PAS reaction (periodic acid-Schiff) for demonstration of neutral muccopolysaccarides. Samples kept in metacrylate were stained toluidine blue and PAS as well. Lipids were detected with toluidine blue in semithin sections. The slides were examined by optical light microscope (Olympus BH 2) and images captured by digital camera.
For TEM small pieces of ovarian tissue were kept in the fluid containing 2.5% glutarealdehyde-phosphate buffer (pH 7.4) at 4oC. The samples were postfixed in 1% OsO4- phosphate buffer on ice. Fixed samples were dehydrated in a series of graded ethanol solutions, then embedded in Epon 812, cut at 500-700Å (Reichert Om U3). Ultrathin sections were double stained with uranyl acetate and lead citrate and examined by use of a JOEL JEM-100C.
In each section, number of the different stages during each month was determined per microscopic field (X100) in the interior, middle and posterior parts of ovary for each individual. Oocytes sectioned through the nucleus were measured using an image analyzer (Media Cybernetics, Silver Spring, MD).
RESULTS
Annual cycle of the ovary: The GSI values were low in June and August (1.6±0.1 and 1.7±0.1%, respectively). In May a little increase was seen as a sign of exogenous vitellogenesis. There was significant increase in October (10.3±1.54%) and it stayed 12.4±1.81% in November. Thereafter, another significant increase was seen in December (18.8±1.2%). The GSI values were suddenly decreased in January and February (1.8± 0.15-0.8±0.1%, respectively). The spawning period is characterized by a high GSI (18.8%) and high oocyte diameter (4466 μm). In December ovulation occur and last until February (Fig. 1).
Light and electron microscopy: The ovary consists of ovigerous lamellae transversally arranged along longitudinal axis. The lamellae contain follicles in various developmental stages (Fig. 2). Somatic prefollicular cells occur in nests in ovigerous lamellae together with oogonia and early oocytes. Oogenesis commences when oogonia are transformed into oocyte. Simultaneously a layer squamous of follicle cells envelops each oocyte (Fig. 3). They are present close to the very young oocytes (perinucleolar stage) initially. The zona radiata starts to arise between follicle cells and oocyte. We observed that follicle cells surrounded by basal lamina (Fig. 4). They are spindle-shaped and have a large nucleus but little cytoplasm. They constituted one layer with granular interior including small droplets. Short microvilli appear in plasma membrane of the oocyte and the apex of the follicular cells (Fig. 5).
With the oocyte growth, the follicle cells underwent changes in their morphology. At the beginning of vitellogenesis they gradually changed in shape from squamous to cuboidal. We determined that the nuclei have prominent nucleoli (Fig. 6).
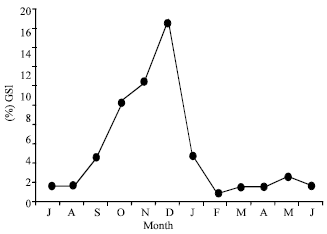 | |
| Fig. 1: | Changes in the gonadosomatic index in the rainbow trout during annual cycle |
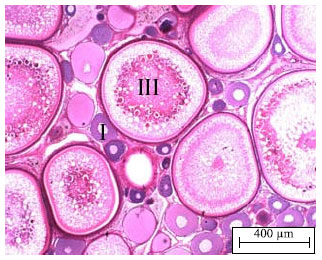 | |
| Fig. 2: | Previtellogenic oocytes (I) and vitellogenic oocytes (III) are seen in the ovary. Dye: PAS-Haemalun |
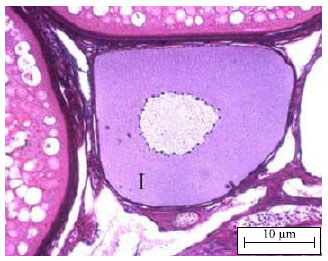 | |
| Fig. 3: | Previtellogenic oocyte (I) surrounded by thin follicle epithelium |
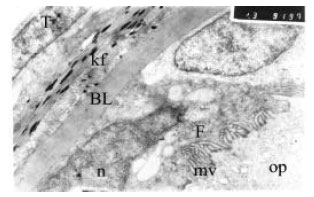 | |
| Fig. 4: | Electron micrograph of primary growth phase oocyte form the ovary of trout. Follicle cells (F) surrounded by basal lamina (bl) show abundant microvilli (mv) projecting from oocyte (O) surface. Thecal cells (T) form the outermost layer of the follicles in the ovary. X16.6k |
 | |
| Fig. 5: | Follicle wall of early vitellogenic oocyte. Theca interna (ti) and theca externe (te) differentiated in this stage. X10k |
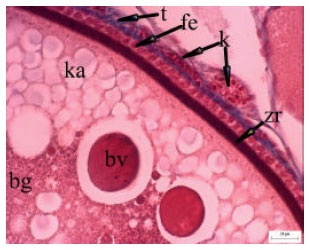 | |
| Fig. 6: | The vitellogenic oocyte covered by cuboidal follicle cells (fe), between zona radiata (zr) and theca (t). Connective tissue, is dyed blue and capillary (k) are showed in the vitellogenic oocytes. Dye: Azan trichrom |
 | |
| Fig. 7: | Follicle cells incluing several mitocondria (m), vesicules (v) and nucleus (n). Basal lamina (bl) and theca (t) surrounding follicle. X25k |
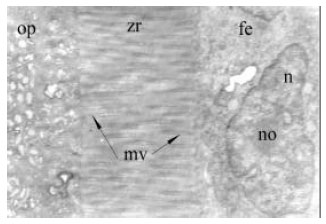 | |
| Fig. 8: | The ooplasma (op) of the vitellogenic oocyte and follicle wall, consist of follicular epithelium (fe), zona radiata (zr) and microvilli (mv). Follicular cell have a ear shape nucleus (n) and its nucleolus (no). X13k |
 | |
| Fig. 9: | The microvilli (mv) extended between the ooplasma (op) and follicle cells (fe). X15k |
 | |
| Fig. 10: | Intercellular space (v) with amorphous materials and heterochromatin (hc) area are seen in the follicular cell. X25k |
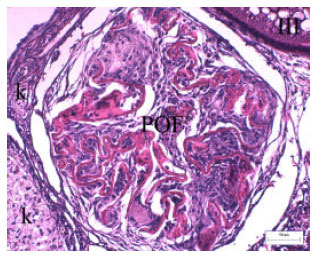 | |
| Fig. 11: | Post ovulatory follicle (POF) and capillary (k) are seen in the ovary. Dye: PAS-Haemalun |
In EM studies have revealed the presence of various cells organelles, including well developed Golgi complex, various vesicules, lipid droplets and mitochondria (Fig. 7). The presence of these elements in the cytoplasm of follicle cells as described above is indicative of protein synthesis (Guraya, 1986). These proteins transport into the oocyte by microvilli (Fig. 8 and 9). The heterochromatin is mainly positioned peripherally in the nuclei of follicle cells. In the later stages of oocyte growth, follicular epithelium was stretched and observed some intercellular spaces. The spaces were filled with amorphous material (Fig. 10). After ovulation we are distinguished that follicle epithelium and theca were folded (Fig. 11).
DISCUSSION
Despite the wide geographic distribution of the trout, the timing of the spawning period does not show any important variation between different areas. Spawning generally take place during early winter. The present observations like to van den Hurk et al. (1979), Tyler et al. (1990) and Tyler (1991).
The oocyte follicular epithelium in teleost fish is derived from prefollicular somatic cells associated with germ cell clusters or nests in the ovigerous lamellae (Grier, 2000; Quagio-Grassiotto and Guimarães, 2003). The prefollicular cells originate from the ovigerous lamellae epithelium (Grier, 2000). In O. mykiss as in the other Teleostei when the oocytes leave the nests, prefollicular cells accompany them and form an epithelium surrounding the oocytes continuously (Wallace and Selman, 1981; Grier, 2000; Quagio-Grassiotto and Guimarães, 2003).
During the primary oocyte growth the follicular epithelium and theca transform. The initially is composed of a layer of squamous follicular cells, derived from the connective tissue forms theca. Basal lamina lies between them (Wallace and Selman, 1981; de Vlaming 1983; Guraya, 1986; Grier, 2000; Quagio-Grassiotto and Guimarães, 2003). During the secondary growth phase or vitellogenesis the follicular epithelium is recognized as protein-producing cells. Basal lamina thickness and the theca interna vascularization like described by Wourms (1976). The intercellular spaces of the follicular epithelium contain material of low electron density (Guraya, 1986; Kjesbu and Kryvi, 1998; Quagio-Grassiotto and Guimarães, 2003). Zona radiata consisted of a thick densely staining only one layer in the trout similar to Larson, 1986. This characteristic is likely to be related to its oviparous nature.
Electron microscopy and histochemical studies have suggested that the follicular cells would be involved in synthesis different proteins during oocyte development. Part of these proteins would be used by the oocyte for its development.
ACKNOWLEDGMENTS
This study was funded within the framework of a research program financed by Marmara University, Sciencitific Researche Found (Project number, FEN-BSE-085/0512019. The authors are indebted to technicians in I.U. Histology and Embryology Laboratory, for their assistance in preparing EM sections.
REFERENCES
- Tyler, C.R. and J.P. Sumpter, 1996. Oocyte growth and development in teleosts. Rev. Fish Biol. Fish., 6: 287-318.
CrossRefDirect Link - Tyler, C.R., J.P. Sumpter and P.R. Witthames, 1990. The dynamics of oocyte growth during vitellogenesis in the rainbow trout (Oncorhynchus mykiss). Biol. Reprod., 43: 202-209.
PubMed - Quagio-Grassiotto, I. and A.C.D. Guimaraes, 2003. Follicular epithelium, theca and egg envelope formation in Serrasalmus spilopleura (Teleostei, characiformes, Characidae). Acta Zool., 282: 121-129.
Direct Link - Krisfalusi, M., P.A. Wheeler, G.H. Thorgaard and J.G. Cloud, 2000. Gonadal morphology of female diploid gynogenetic and triploid rainbow trout. J. Exp. Zool., 286: 505-512.
Direct Link