
Gamalat A. Abd-Elaah
Department of Botany, Faculty of Science, South Valley University, Sohag, Egypt
A.A. Galal
Department of Botany, Faculty of Science, South Valley University, Sohag, Egypt
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 5 | Page No.: 898-903







ABSTRACT
Thirteen species belonging to six genera of zoosporic fungi, yielding 356 fungal colonies were isolated from 30 soil samples collected from Sohag city. The effect of different levels (0.0, 50.0, 100.0, 150.0 and 300.0 ppm) of the fungicide benomyl on the morphological structures of mycelial, zoosporangia, sexual organs and gemmae of Pythium aphanidermatum and Achlya americana was studied on malt extract medium and callus cultures of garlic. Steroles was determined in garlic callus cultures treated with different levels of benomyl fungicide. GLC analysis showed differences in sterol contents between treated calli of garlic and control ones, which did not meet Achlya americana and Pythium aphanidermatum fungi requirements.
PDF Abstract XML References Citation
How to cite this article
Gamalat A. Abd-Elaah and A.A. Galal, 2006. Effect of Benomyl Treated Garlic on Growth and Sporulation of Pythium aphanidermatum and Achlya americana. Pakistan Journal of Biological Sciences, 9: 898-903.
DOI: 10.3923/pjbs.2006.898.903
URL: https://scialert.net/abstract/?doi=pjbs.2006.898.903
DOI: 10.3923/pjbs.2006.898.903
URL: https://scialert.net/abstract/?doi=pjbs.2006.898.903
INTRODUCTION
Fungicides may have a direct impact on the natural occurrence, infectivity and population dynamics of entomopathogenic fungi. They also affect other macro- and micro-organisms in the soil which may interact with the entomopathogenic fungi. Benomyl is among the most frequently used systemic fungicides against pathogenic fungi of several plant species including garlic (Anonymous, 1997). The primary mode of action of benomyl has been identified as the specific binding to the β-tubulin subunit of fungal tubulin and consequently an interference with microtubules assembly (Hammersachlag and Sisler, 1973; Howard and Aist, 1980; Issac, 1992). Microtubules are major components of the fungal cytoskeleton and are involved in meiosis and mitosis both of which are blocked in the presence of benzimidazole benomyl (Chiocchio et al., 2000; Corbett et al., 1984; Davidse, 1986, 1987).
The effect of sterols on the vegetative growth is thought to be the result of membrane stabilization and on increase the rate of hyphal growth (Nes, 1974, 1987). Hendrix (1970) and Nes (1987) reported that the addition of sterol to basal media stimulate the vegetative growth and reproduction of Phytophthora sp. and Pythium sp. The dramatic enhancement of growth and reproduction following the addition of low levels of sterols after a period of sterol-less growth has been observed (Nes, 1987; Weete, 1989; Elliot, 1977).
Oomycetes are unable to synthesize sterols de novo from mevalonate, but they possess remnants of the sterol biosynthetic pathway which enable them to carry out certain transformation when supplied with exogenous sterols (Nes, 1987; Elliot, 1977). It was found that deficiency in exogenous sterols in the diet of fungi affect the growth and sporulation process (Elliot, 1977; Lutova et al., 1994; Koller, 1991). The present investigation aimed to study the effect of growth inhibiting fungicide (benzimidazole benomyl) treated plant on the growth and sporulation process in Pythium aphanidermatum and Achlya americana.
MATERIALS AND METHODS
Isolation of zoosporic fungi from soil: Thirty soil samples (ca. 500 g) were collected from soils cultivated with wheat, onion, garlic and alfalfa at Sohag Governorate in clean plastic bags and brought to the laboratory. The soil temperature and the pH values of the soil were determined in situation. For the recovery of zoosporic fungi from soil samples, the baiting technique using sesame seeds was used. Soil sample (5 g) were introduced into a sterile, clean, 12 cm petri dish (6 replicates) containing 20 mL of sterile distilled water and 10-12 sterilized sesame seeds. The seeded plates were incubated at 20°C for 3-6 weeks during which the growing fungi were examined, purified (Willoughby and Pickering, 1977) and counted. The identification of fungal genera and species was performed after (Coker, 1923; Fitzpatrick, 1930; Johnson, 1956). Five replicates were used for each sample. For the determination of fungal population, the zoosporic fungal species appearing on one plate was counted as one colony.
Effect of benomyl on Pythium aphanidermatum and Achlya americana: The effect of the fungicide benomyl [methyl 1-(butylcarbamoyl)-2-benzaimidazol-ecarbamate, C14H18N4O3, Benlate, 50% active ingredient) on the morphological structures of Pythium aphanidermatum and Achlya americana was studied on malt extract liquid medium.
Treatment of fungi with fungicide on malt extract medium: Malt extract liquid medium (pH 6) supplemented with five different concentration (0, 50, 100, 150 and 300 ppm) of the fungicide benomyl were inoculated with zoospore suspensions of the aquatic fungal species Pythium aphanidermatum and Achlya americana. Five replicates including 5 plates replicate-1 were used for each concentration. The cultures microscopically examined for mycelial growth and the formation of zoosporangia, sexual organs and gemmae after 2, 4, 6 and 8 days of incubation at 20°C.
Callus cultures as a source of exogenous sterols: Mature bulbs of the Egyptian garlic (Allium sativum L.) cultivar Balady were kept at 5°C for 2 months. Then, the cloves were dehisced and surface-sterilized by rinsing in 70% alcohol for 1 min followed by immersion in 20% (v/v) commercial bleach (which contained 5.5% NaOCl) for 30 min. They were then washed four times in sterile distilled water and germinated on half strength hormone-free MS (Murashige and Skoog, 1962) medium. Young leaf parts inside the cloves were aseptically isolated and served as explants. The leaf explants were then cultured on the callus induction medium. This medium was based on BDS medium (Dunstan and Short, 1977) and supplemented with 1.0 mg L-1 2, 4-Dichlorophenoxyacetic acid, 2.0 mg L-1 kinetin, 100 mg L-1 myo-inositol and 30.0 g L-1 sucrose. The medium was solidified with 0.8% agar (pH was 5.5) and autoclaved for 15 min at 1.2 kg cm-2. The cultures were incubated in continuous illumination at 25°C. The developed calli were subcultured onto the same medium every three weeks.
Callus pieces (ca 1.0 g each) were transferred to petri dish (10 cm) containing callus induction liquid medium for 3 weeks supplemented with 0.0, 50.0, 100.0, 150.0 or 300.0 ppm of the fungicide benomyl. Then callus cultures were inoculated with zoospore suspensions of Pythium aphanidermatum and Achlya americana. The experiment was replicated three times using 4 callus pieces per each replicate and incubated in continuous illumination at 25°C. The hyphal growth and the formation of zoosporangia and Gemmae were examined after 2, 4, 6 and 8 days of incubation.
Determination of sterol: Treated calli with 100 ppm benomyl from 8 days cultured were subjected for sterol determination following. Grinned dried sample (ca. 0.5 g) was accurately weighed then extracted in a soxhlet apparatus with petroleum ether (40-60°C) for several times.
The solvent was removed by evaporation under reduced pressure and the percentage of total lipids was calculated. The total lipid were separated and fractionated according to the method of Farag et al. (1986). For separating fatty acids and unsaponifiobles, lipid materials were saponified with methalonic KOH (20% w/v) for 24 h at room temperature the unsponifiables were extracted three times with diethyl ether (peroxide free). The aqueous layer was acidified by hydrochloric acids (20% v/v) and the liberated fatty acids were extracted times with peroxide free diethyl ether. The combined extracts of unsaponifiable and fatty acids were washed several times with distilled water until the washing was neutral, then dried with anhydrous sodium sulphate. The unsaponifiables were also fractionated on Gas Liquid Chromatography (GLC) using cooled glass column (2.8 mx. mm) packed with Diatomite-e (100-120 mesh) and coated with 1% or-17 oven temperature was programmed at 10°C –1 min from 70 to 270°C then so thermally at 270°C for 25 min and nitrogen flow rate was 30 mL min-1. Injector and detector temperatures were maintained at 300°C.
RESULTS AND DISCUSSION
Thirteen species belonging to six genera of zoosporic fungi yielding 356 fungal colonies were isolated from 30 soil samples collected from Sohag city (Table 1). These samples were cultivated with wheat, onion, garlic and alfalfa. The dominant species were Allomyces (2 species), Saprolegnia (4 species), Achlya (3 species) and Pythium (2 species). Isoachlya eccentrica and Dictyuchus sterilis were represented by moderate and low occurrence, respectively.
The occurrence and distribution of aquatic fungi were investigated by El-Hissy and Abd-Elaah (1989) in 315 soil samples collected from all over Upper Egypt from Aswan to El-Giza including El-Fayoum Governorate.
| Table 1: | Frequency of occurrence of aquatic fungi recovered from 30 cultivated soil samples collected from Sohag Governorate |
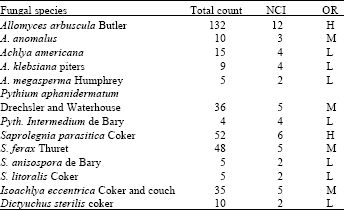 | |
TC: Total Count, H: High occurrence more than 10 cases, M: Moderate occurrence; between 5-10 cases, L: Low occurrence; between 2-5 cases | |
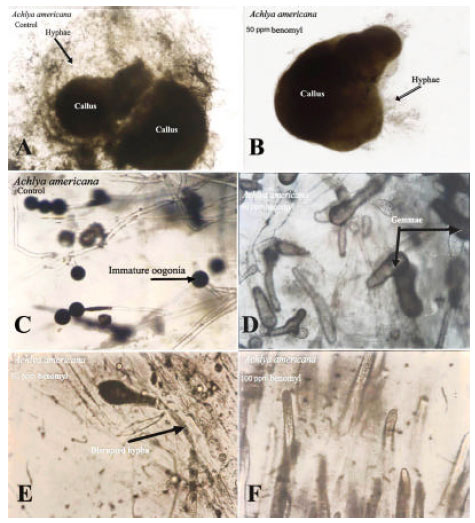 | |
| Fig. 1: | Growing of Achlya americana on control and benomyl treated cultures: (A): Growing hyphae on garlic callus pieces in control medium. (B): Low growth of hyphae on garlic callus treated with 50 ppm benomyl. (C): Immature oogonia on control medium. (D) Gemmae on 50 ppm benomyl. (E): Disrupted hyphae on 50 ppm benomyl. (F): The growing hyphae at 100 ppm benomyl |
Hassan and Fadl-Allah (1992) studied some zoosporic fungi in soils of Upper Egypt. However, Abd-Elaah (1998) identified nineteen species recovered from 14 soil samples collected from different sites of the Red Sea Governorate. Furthermore, Abd-Elaah (1999) isolated 36 species in addition to three unidentified ones, which belong to eighteen fungal genera 60 soil samples collected seasonally from the cultivated clay soil at Sohag. She isolated Allomyces, Achlya, Pythium, Saprolegnia, Dictyuchus and Isoachlya, results found that the highest number of aquatic fungal species was recovered during winter.
Effect of benomyl on Pythium aphanidermatum and Achlya americana: The effect of different levels (0.0, 50.0, 100.0, 150.0 and 300.0 ppm) of the fungicide benomyl on the morphological structures of mycelial, zoosporangia, sexual organs and gemmae of Pythium aphanidermatum and Achlya americana was studied on malt extract medium and callus cultures of garlic.
In malt extract medium, the density of vegetative mycelia of Pythium aphanidermatum and Achlya americana increased with the increase of the incubation period in case of the control (0.0 benomyl) treatment. While at 50 ppm of benomyl, low density of mycelia was observed in both tested fungi after 4 days of incubation this agree with the results obtained by Gangawane and Shaikh (1988). No mycelial growth appeared at the concentrations exceeded 50 ppm of the fungicide whatever the period of incubation used. High number of zoosporangia was formed when Achlya americana and Pythium aphanidermatum cultured on the control medium and incubated at 20°C for 8 days. While, low number of zoosporangia was observed after 8 days at 50 ppm of the fungicide benomyl.
| Table 2: | Effect of benzimidazole fungicide benomyl on the free sterol contents of garlic callus |
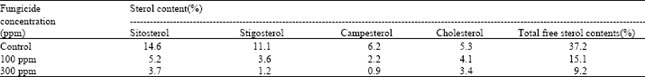 | |
| % was calculated relative to the total percentage of unsaponifiable matters | |
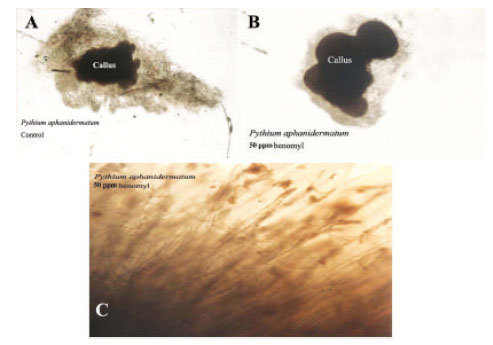 | |
| Fig. 2: | Growing of Pythium aphanidermatum on control and benomyl treated cultures: (A): Growing hyphae on garlic callus pieces in control medium. (B): Low growth of hyphae on garlic callus pieces treated with 50 ppm benomyl. (C): The growing hyphae at 50 ppm benomyl |
The results also showed that the number of the discharged zoosporangia was high at control treatment but rare at 50 ppm of the fungicide which in agreement with those obtained by Suryanarayna and Thomas (1986). The fungicide benomyl completely inhibited the formation of oogonia and antheridia at any concentration used. The oogonia and antheridia were observed after 7 days of incubation in the control of the two tested fungi. Gemmae were observed in cultures free from the fungicide and at 50 ppm concentration of the fungicide.
The effect of treated garlic callus culture with different concentrations of the fungicide benomyl (0.0, 50.0, 100.0, 150.0 and 300.0 ppm) on Pythium aphanidermatum and Achlya americana revealed normal growth of the tested fungi on the control medium (0.0 benomyl). The tested fungi grew slowly on the treated calli with concentration 50 ppm and formed gemmae while no sporangia were observed (Fig. 1 and 2). At concentration 100 ppm the hyphal growth was sharply affected and a little number of hyphae and gemmae were observed.
GLC analysis of free sterol contents of garlic callus treated with two different concentrations (100 and 300 ppm) of benzimidiazole fungicide benomyl, showed that the percentage of free sterol contents decrease as well as the concentration of fungicide benomyl increase compared with control ones (Table 2). There was a dramatic quantitative reduction with the increase of fungicide concentration. Also the total percentage of free sterol contents decreased as well as the concentration of fungicide increased.
The obtained results were in agreement with those obtained by Hippe (1991) who reported that the fungicide acylalanines inhibited the rRNA synthesis and the fungicide carboxamides inhibited the respiration processes. The hyphal growth of the two tested fungi was completely inhibited on the treated calli with higher concentrations (150 ppm and 300 ppm). Similar results were also observed by Hammersachlag and Sisler (1973), Howard and Aist (1980); Issac (1992) and Chiocchio et al. (2000) who showed that the concentrations 0.1 and 0.001 μg mL of benomyl did not inhibit the germination of Glomus mosseae spores, but inhibited the hyphal length of the germ tube.
Also, many explanations have been given by many investigations regarding the inhibition of growth and reproduction of Pythium and Achlya that cultured with the treated calli with the fungicide benomyl. Davidse and Flach (1977) attributed this inhibition to its effect on tubulin integrity. Microtubules are alternating helices of α-and β-tubulins which form an essential part of the cytoskeleton and are active in spindle formation and the segregation of chromosomes in cell division. In this respect, Hammersachlag and Sisler (1973) reported that carbendazim, the common active principle of benomyl disrupt mitosis during cell division at metaphase. The mitotic spindle is distorted and daughter nuclei fail to separate. These results are in agreement with those obtained in the present study. The obtained results indicated that sporulation process in both Pythium and Achlya that grow on the treated calli with benomyl was lower than those of control ones. This may be attributed to the deficiency in sterol contents of calli. This result is in agreement with those obtained by Hendrix (1970), Elliot (1977) and Lutova et al. (1994), who reported that deficiency in exogenous sterols in the diet of fungi affects the sporulation process of fungi. Whereas, the Oomycetes metabolized cycloartenol, the first intermediate in non-photosynthetic organisms which responsible to synthesis it, in preference to lanosterol the first intermediate in non-photosynthetic organisms (Nes, 1987; 1984; Warner and Domnas, 1981; Warner et al., 1983) or this may be attributed to the fungicidal toxicity resulted in accumulation of the fungicide in the treated calli.
REFERENCES
- Abd-Elaah, G.A., 1998. The occurrence of fungi along the red sea coast and variability among isolates of Fusarium as revealed by isozyme analysis. J. Basic Microbiol., 38: 303-311.
CrossRefDirect Link - Abd-Elaah, G.A., 1999. Seasonal variations of zoosporic and terrestrial fungi in cultivated soil at Sohag Governorate and isozyme differences among Achlya species. Afr. J. Mycol. Biotechnol., 7: 7-23.
Direct Link - Chiocchio, V., N. Venedikian, A.E. Martinez, A. Menendez, J.A. Ocampo and A. Godeas, 2000. Effect of the fungicide benomyl on spore germination and hyphal length of the arbuscular mycorrhizal fungus Glomus mosseae. Int. Microbiol., 3: 173-175.
PubMedDirect Link - Davidse, L.G. and W. Flach, 1977. Differential binding of methyl benzimidazol-2-yl carbamate to fungal tubulin as a mechanism of resistance to this antimitotic agent in mutant strains of Aspergillus nidulans. Cell. Biol., 72: 174-193.
PubMedDirect Link - Dunstan, D.I. and K.C. Short, 1977. Improved growth of tissue cultures of the onion, Allium cepa. Physiol. Plant., 41: 70-72.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Davidse, L.C., 1986. Benzimidazole fungicides: Mechanism of action and biological impact. Ann. Rev. Phytopathol., 24: 43-65.
Direct Link