
M. Amiri Andi
Science and Research Campus, Islamic Azad University, Tehran, Iran
M. Shivazad
Department of Animal Sciences, Faculty of Agriculture, Tehran University, Tehran, Iran
S.A. Pourbakhsh
Department of Poultry Diseases Razi Institute, Karaj, Iran
M. Afshar
Researches Center of Agriculture and National Resources, Tehran, Iran
H. Rokni
Researches Center of Agriculture and National Resources, Tehran, Iran
N.E. Shiri
Researches Center of Agriculture and National Resources, Tehran, Iran
A. Mohammadi
Researches Center of Agriculture and National Resources, Tehran, Iran
Z. Salahi
Department of Poultry Diseases Razi Institute, Karaj, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 5 | Page No.: 789-794







ABSTRACT
Broiler breeders were allocated at random to one of seven experimental diets containing graded levels of supplementary vitamin E (0, 20, 40, 60, 80, 100 and 120 IU kg-1 diet) between 26-35 weeks of age. Egg production, egg weight, fertility, livability and hatchability (30 and 35 weeks of age) were not influenced by the level of vitamin E in the breeder diet. But birds receiving 40 IU of vitamin E kg-1 diet had higher hatchability of fertile eggs compared to 0 and 20 IU of vitamin E kg-1 groups at 35 weeks of age (90.79 vs. 85.09 and 86.60%, respectively). Dietary levels of vitamin E did not affect antibody titer (IgG) against Newcastle disease virus at 35 weeks of age. Antibody titer of day old chicks from hens receiving 60 IU of vitamin E kg-1 were higher (p≤0.05) than chicks from hens fed 0 and 20 IU of vitamin E. At 35 weeks of age (not at 31 weeks of age), Hugh units of eggs were higher (p≤0.05) in hens fed 40, 60, 80, 100 and 120 IU of vitamin E kg-1 compared to 0 IU of vitamin E kg-1 (87.06, 92.21, 93.89, 93.25, 94.61 and 93.09, respectively). Age had negative effect on Hugh units of eggs (p≤0.01). The results of this experiment suggest that the vitamin E requirement of broiler breeders for persisting of hatchability may be 40 IU kg-1 and for maximizing passive transfer of antibody from breeder to progeny may be higher.
PDF Abstract XML References Citation
How to cite this article
M. Amiri Andi, M. Shivazad, S.A. Pourbakhsh, M. Afshar, H. Rokni, N.E. Shiri, A. Mohammadi and Z. Salahi, 2006. Effects of Vitamin E in Broiler Breeder Diet on Hatchability, Egg Quality and Breeder and Day Old Chick Immunity. Pakistan Journal of Biological Sciences, 9: 789-794.
DOI: 10.3923/pjbs.2006.789.794
URL: https://scialert.net/abstract/?doi=pjbs.2006.789.794
DOI: 10.3923/pjbs.2006.789.794
URL: https://scialert.net/abstract/?doi=pjbs.2006.789.794
INTRODUCTION
Vitamin E (VE) is a term used to describe two compounds: tocopherols and tocotrienols. These can be found naturally, with the richest sources being vegetable oils, eggs, liver, legumes and in general, green plants. Vitamin E has a number of different but related functions. One of the most important functions is its role as an intracellular antioxidant (Tappel, 1970). Vitamin E has been reported as an excellent biological chain-breaking antioxidant that protects cells and tissue from lipoperoxidative damage induced by free radicals (Yu, 1994).
The age and production stage of a hen affect egg shell structure (Pappas et al., 2005). On the other hand, in storage period, loss of egg internal water and CO2 from the shell pores alter the pH of the albumen (Decuypere et al., 2001). Such changes may also contribute to the reduction in hatchability (Etches, 1996). But no recent data on the effect of VE on egg quality of broiler breeders are available.
A survey of the literature suggests a positive effect of VE on immune functions in poultry (Gore and Qureshi, 1997; Haq et al., 1996; Hossain et al., 1998; Puthpongsiriporn et al., 2001). Vitamin E has been reported to protect cells involved in immune response, such as lymphocytes, macrophages and plasma cells, against oxidative damage and to enhance the function of these cells (Konjufca et al., 2004; Meydani and Beharka, 1998) and high levels of VE increases antibody production (Gore and Qureshi, 1997; Haq et al., 1996; Hossain et al., 1998). Jackson et al. (1978) and Haq et al. (1996) demonstrated high levels of VE in breeder diet affect serum antibody titer of progeny against antigenic stimulation of Brucella abortus and Newcastle Disease Virus (NDV). In addition, it is generally believed that nutrient levels considered adequate for growth may not be adequate for optimal immune response and disease resistance (Nockels, 1988). On the other hand, recent data (Hossain et al., 1998) were shown that for determining of broiler breeder requirements of VE we have to measure not only breeder immunity and hatchability, but also progeny immunity. In spite of above explanations, NRC (1994) was not specified VE levels for broiler breeders.
The present study was part of a project designed to assess the effect of different levels of VE in broiler breeder diet on hatchability, egg quality, immunity (maternal and neonatal) and progeny performance. The objectives of the study in this part were to evaluate hatchability, egg quality, breeder immunity and antibody transfer to the progeny.
MATERIALS AND METHODS
Housing and management: This study was conducted in Khojir Station, Researches Center of Agriculture and Natural Resources, Tehran Province, Iran (started at 03/07/2005 and finished at 12/03/2005). Five hundred female and 50 male day old Arian broiler breeders were randomly allocated to one of 28 pens (2.2x2.0 m) and pullets and cockerels were reared separately in groups of 30 and 25, respectively. The birds were reared on a restricted feeding program recommended by the Arian broiler breeder’s management guide. At 20 weeks of age all the birds were weighed and 280 females and 28 males were selected within ±40 g of the mean body weight and were distributed randomly to one of 28 pens with 10 females and one male per pen. Each pen was equipped with one tube male exclusion feeder for females and one tube feeder for male mounted out of reach of the females, an automatic water and 2-hole nest-box. Light stimulation started at 21 weeks of age with 9 h photoperiods and gradually increased till 27 weeks of age and fixed (16 h). The birds were weighed at the end of the experiment (35 weeks of age).
Treatments: The seven experimental diets (Table 1) were provided when breeder hens were at 26 weeks of age. The experimental diets for females were based on corn and soybean meal (with barley, 6.98%) and formulated to contain 16.5 g kg-1 CP and 2700 Kcal ME kg-1 with seven levels of VE ( 0, 20, 40, 60, 80, 100 and 120 IU kg-1). They were each offered to four replicate groups in a completely randomized design. From 20 weeks of age all males were fed a diet containing 13 g kg CP and 2800 Kcal kg ME. Daily feed allocation per female was based on production rate and body weight whereas for the male it was based on body weight according to the management guide. Alpha tocopherol content of the experimental diets analysed by HPLC procedure (laboratories of Martin-Luther-University, Nutrition Science Institute, Emil-Abderhalden Street 26, 06108, Halle/s, Germany).
| Table 1: | Composition of experimental diets |
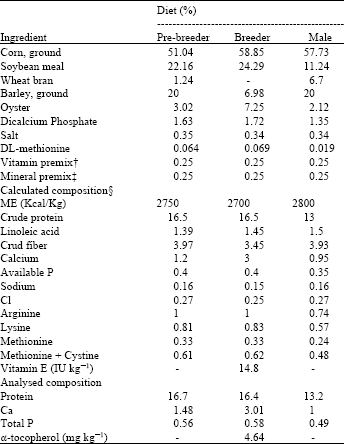 | |
†Vitamin premix for breeder females contained no Vitamin E (20 IU Vitamin E, for male). Other vitamins provided per kg of diet: Vitamin A, 9500 IU; Vitamin D3, 2000 IU; Vitamin K3, 2.5 mg; Thiamin, 1.97 mg; Riboflavin, 6 mg; Niacin, 25 mg; Pantothenic acid, 12 mg; Biotin, 0.1 mg; Folic acid, 0.8 mg; Vitamin B12, 14 μg and Ethoxyquin, 100 mg. Vitamin E (dl-α-tocopherol acetate ) were added to the basal diet according to the treatment, ‡ Mineral premix for breeder females provided per kg of diet: Manganese, 99.2 mg; Zinc, 60 mg; Iron, 80 mg; Copper, 6 mg; Iodine, 0.868 mg and selenium, 0.2 mg, § On the basis of NRC (1994) ingredients analysis | |
Egg production and egg quality: Eggs were collected and recorded daily and egg production calculated weekly. Hatchable eggs were stored at 15-16°C until setting in the incubator. Mean egg weight per pen was determined by weighing all the eggs on three consecutive days every week. The yolk, albumen and shell weight were determined using 8 eggs per treatment at 31 and 35 weeks of age. Albumen heights were measured by an albumen height determiner (OSK-13471, OGAWA SEIKI CO., LTD. JAPAN). The Hugh unit of the albumen was calculated by using the Hugh unit formula (Eisen et al., 1962). Shell thicknesses of these eggs were measured by a micrometer (OSK-13469, OGAWA SEIKI CO., LTD. JAPAN) and albumen pHs were determined by a pH meter (Model EC20 pH/ISE Meter, HACH COMPANY, USA).
Fertility, hatchability and chick quality: Every two weeks 10 eggs from each pen were collected randomly, identified and incubated in an incubator with temperature 37.5°C and 60% relative humidity. After 3 days of incubation all eggs were opened to determine the fertility. To determine the hatchability and average chick weight a total of 36 eggs per replicate group was collected at 30 weeks age and 72 eggs per replicate group at 35 weeks of age. They were identified, weighed and set in an incubator (JAMESWAY incubator) for 19 days, after which they were transferred to a hatcher until hatched. On the 14th day the eggs were candled to determine the number of infertile eggs and early dead embryos. After 21 days incubation, the number and weights of live chicks were recorded. Hatchability of the eggs from each replicate pen was calculated and expressed relative to the total of eggs set and to the fertile eggs.
Serum antibody titer: At the end of 26 weeks of age, before feeding of the experimental diets, blood specimens were collected from individual birds by brachial venipuncture. Blood samples were allowed to clot at ambient temperature, centrifuged for 10 min at 3,000 rpm and serum from each sample was extracted, using individual sterile Pasture pipets. Serums of hens of one pen were pooled. The pooled serum samples were kept frozen until antibody titers against NDV were measured. Antibody titers (IgG) were assessed by ELISA method using the Newcastle disease antibody test kit (ProFLOK®PLUS, SYNBIOTICS CORPORATION, USA). Nine weeks after the initiation of experimental feeding (at 35 weeks of age), blood specimens were collected from all breeders and sera extracted using procedures. Blood samples were collected from 8 chicks (day old) of each treatment group (second incubation). Sera extraction (not pooled) and antibody titers were done similar to the procedure described above.
Statistical analysis: The data were analyzed by ANOVA in a one-way classification using the General Linear Models (GLM) procedure of the SAS Institute (SAS Institute, 1996). Means were separated using the Duncan's multiple range test option.
RESULTS
Alpha tocopherol contents of the experimental diets were 4.64, 23.64, 40.1, 54.3, 72.91, 98.91 and 110.8 mg kg-1 diet, according to the supplemental VE.
Control treatment (0 IU of VE kg-1 diet) had the lowest egg production and broiler breeders fed 80 IU of VE kg-1 diet had the highest (77.9 and 86.5%, respectively) (Table 2) Vitamin E level in broiler breeder diet did not affect hatchability (Table 3). But, the control had the lowest hatchability in the second incubation (85.09% of fertile eggs) and hens fed 40 IU of VE kg-1 had the greatest one (90.79% of fertile eggs). In addition, the level of 20 IU of VE kg-1 diet did not prevent the decline of hatchability in the 2nd incubation (86.60% of fertile eggs). Birds at 30 weeks of age had higher pooled hatchability than birds at 35 weeks of age (90.12 vs. 88.39% of fertile eggs, respectively). Different levels of VE in broiler breeder diet did not affect chick weight and ratio of chick weight to egg weight (data not shown).
| Table 2: | Effects of dietary Vitamin E on egg production, egg weight, fertility and body weight changes and livability of breeders between 27-35 weeks of age1 |
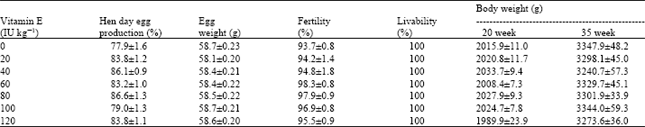 | |
| Values are means±SEM | |
| Table 3: | Effects of Vitamin E in broiler breeder diet on hatchability1 |
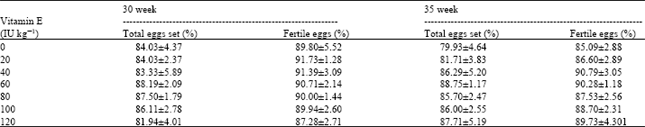 | |
| Values are means±SEM | |
| Table 4: | Effects of Vitamin E in broiler breeder diet on antibody (IgG) titer of breeders and on passive antibody titer transport to progeny (day old chicks)1 |
 | |
| 1.Values are means±SEM and log10 titer, a-b.Values in the same column with no common superscript differ significantly (p≤0.05) | |
| Table 5: | Effects of Vitamin E in breeder diet on egg quality1 |
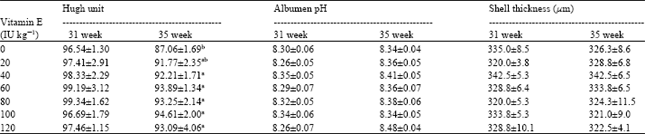 | |
| 1.Values are means±SEM, a-b.Values in the same column with no common superscript differ significantly (p≤0.05) | |
Antibody titers of hens did not differ after nine weeks of feeding of different VE levels in their diet. But, hens receiving 120 IU of VE kg-1 diet had the greatest antibody titer and the control had the lowest one at 35 weeks of age (4.10 and 3.91, respectively). Chicks from hens receiving 60 IU of VE kg-1 diet had the highest and significant antibody titer (p≤0.05) compared to 0 and 20 IU of VE kg-1 groups (3.98 vs. 3.76 and 3.75, respectively). Parents transferred 97% antibody titer to the progeny (Table 4).
Table 5 shows the Hugh unit of the albumen, albumen pH and shell thickness of eggs at 31 and 35 weeks of age. At 31 weeks of age, there were no significant differences in Hugh units among treatments, but the control group had the lowest (96.54) and birds fed 80 IU of VE kg-1 had the highest albumen Hugh unit (99.34). In the second measurement (at 35 weeks of age), Hugh units were significantly higher (p≤0.05) for birds fed 40, 60, 80, 100 and 120 IU of VE kg-1 diet compared to the control (92.21, 93.89, 93.25, 94.61 and 93.09 vs. 87.06, respectively). Pooled Hugh unit at 35 weeks of age decreased (p≤0.01) by 5.23 unit compared to 31 weeks of age (97.79 vs. 92.56). Different level of dietary VE did not influence albumen pH at 31 and 35 weeks of age, but, hens had higher (p≤0.01) pooled albumen pH at 35 \weeks of age compared to another week (8.38 vs. 8.30). Shell thicknesses were not affected by supplemental VE in hen's diet. Egg content weights (and percentages) were not influenced by treatments (data not shown).
DISCUSSION
Vitamin E is essential for normal hatchability (Kling and Soares, 1980). The VE requirement for chickens is usually estimated on the basis of hatching and egg production (NRC, 1971), but if other functions (such as immune responses) are also considered, this requirement may change (Tengerdy and Nockels, 1973). On the other hand, NRC (1994) has not specified the VE levels for meat type breeders but for egg type breeders with a daily feed intake of 100 g/hen/day the recommended level is 10 mg kg-1 diet. We concluded VE level of 40 IU kg-1 in Arian broiler breeder diet is the best level for egg production and persisting of hatchability. Data of hatchability are compatible with the work of Hossain et al. (1998) who used 25, 50, 75 and 100 mg of VE kg-1 in Ross broiler breeder diet and showed that breeders receiving 50 mg of VE kg-1 had the highest hatchability at 40 weeks of age. Leeson and Summers (1991 and 1997) recommended 20 and 25 mg of VE kg-1 diet for broiler breeders. But, it seems new broiler breeders need VE higher than 20 IU kg-1 diet. Therefore, VE level of 40 IU kg-1 in Arian broiler breeder diet for persisting of hatchability is recommendable.
In this study we concluded the level of VE higher than 20 IU kg-1 diet results in higher egg internal quality (Hugh unit) at 35 weeks of age. Similar results were reported by Puthpongsiriporn et al. (2001) in laying hens. In their experiment, Hugh units were higher in hens fed 65 IU of VE kg-1 compared to 25 and 45 IU kg-1. Vitamin E in maternal diet enhances antioxidant status of egg (Surai, 2000) and protects proteins and lipids of egg from oxidation (Puthpongsiriporn et al., 2001), so had beneficial effect on Hugh unit and probably subsequent hatchability. On the other hand, in our experiment the younger breeders had the higher Hugh unit of albumen. It was reported that the high ovomucin content of the albumen in eggs from younger birds may be associated with a relatively high Hugh unit (Pappas et al., 2005). Our observation is in line with previous reports noting that Hugh unit is influenced by the age of the hen (Brake et al., 1997; Lapao et al., 1999). It seems there is a relationship between albumen Hugh unit and hatchability. This finding is compatible with the opinion of Etches (1996). The birds had higher pH at 35 weeks of age than 31 weeks of age. The higher pH (at 35 weeks of age) results in dissociation of the albumen's proteins in particular (lysozyme and ovomucin), which in turn reduces viscosity of the albumn (Powrie, 1977) and subsequent decline in Hugh unit. No recent data on the effect of supplemental VE in breeder diet on egg quality are available. We recommend further research in this field.
Present experiment showed no effect of supplemental VE on broiler breeders' antibody (IgG) titer against NDV. Similar results were also reported by Haq et al. (1996), Hossain et al. (1998) and Jackson et al. (1978). Conversely, chicks from hens supplemented with 60 IU of VE had significantly higher antibody titer at 1 day of age than chicks from the control and 20 IU of VE kg-1 group. Jackson et al. (1978) attributed this phenomenon to selective antibody absorption by ovarian tissue. In addition, Jackson et al. (1978) and Nockels (1979) have shown that passively transferred antibody levels were significantly increased in the plasma of 2-and 7-d-old chicks when hens were fed high levels of VE in the diet. Babinszky et al. (1991) reported increased antibody levels in serum of 1-wk-old piglets when VE was supplemented to maternal diet. Higher maternal antibody transfer to progeny with supplemental VE in the broiler breeder diet means higher humoral immunity in day old chicks and subsequent disease resistance.
The results of this research suggest that the VE requirement of broiler breeders for persisting of hatchability may be 40 IU kg-1 and for maximizing the immune responses of progeny and egg albumen Hugh unit may be higher. However, further research needs to be done to establish the most appropriate dietary concentration of VE for broiler breeders.
ACKNOWLEDGMENTS
The authors thank Research Center of Agriculture and Natural Resources of Tehran Province for supplying of research costs and equipments.
REFERENCES
- Babinszky, L., D.J. Langhout, M.W.A. Verstegen, L.A. Den Hartog, P. Joling and M. Nieuwland, 1991. Effect of vitamin E and fat source in sows' diets on immune response of suckling and weaned piglets. J. Anim. Sci., 69: 1833-1842.
Direct Link - Decuypere, E., K. Tona, V. Bruggema and F. Bamelis, 2001. The day-old chick: A crucial hinge between breeders and broilers. World's Poult. Sci. J., 57: 127-139.
Direct Link - Eisen, E.J., B.B. Bohren and H.E. McKean, 1962. The Haugh unit as a measure of egg albumen quality. Poult. Sci., 41: 1461-1468.
CrossRefDirect Link - Gore, A.B. and M.A. Qureshi, 1997. Enhancement of humoral and cellular immunity by vitamin E after embryonic exposure. Poult. Sci., 76: 984-991.
Direct Link - Hossain, S.M., S.L. Barreto, A.G. Bertechini, A.M. Rios and C.G. Silva, 1998. Influence of dietary Vitamin E level on egg production of broiler breeders and on the growth and immune response of progeny in comparison with the progeny from eggs injected with Vitamin E. Anim. Feed Sci. Technol., 73: 307-317.
Direct Link - Konjufca, V.K., W.G. Bottje, T.K. Bersi and G.F. Erf, 2004. Influence of dietary vitamin E on phagocytic functions of macrophages in broilers. Poult. Sci., 83: 1530-1534.
PubMed - Lapao, C., L.T. Gama and M.C. Soares, 1999. Effects of broiler breeder age and length of egg storage on albumen characteristics and hatchability. Poult. Sci., 78: 640-645.
Direct Link - Meydani, S.N. and A.A. Beharka, 1998. Recent developments in vitamin E and immune response. Nutr. Rev., 56: S49-58.
Direct Link - Nockels, C.F., 1979. Protective effects of supplemental vitamin E against infection. Fed. Proc., 38: 2134-2138.
Direct Link - NRC., 1994. Nutrient Requirements of Poultry. 9th Edn., National Academy Press, Washington, DC., USA., ISBN-13: 9780309048927, Pages: 176.
Direct Link - Puthpongsiriporn, U., S.E. Scheideler, J.L. Sell and M.M. Beck, 2001. Effects of vitamin E and C supplementation on performance, In vitro lymphocyte proliferation and antioxidant status of laying hens during heat stress. Poult. Sci., 80: 1190-1200.
CrossRefPubMedDirect Link - Surai, P.F., 2000. Effect of selenium and vitamin E content of the maternal diet on the antioxidant system of the yolk and the developing chick. B. Poult. Sci., 41: 235-243.
CrossRefDirect Link - Tappel, A.L., 1970. Biological antioxidant protection against lipid peroxide damage. Am. J. Clin. Nutr., 23: 1137-1139.
Direct Link - Tengerdy, R.P. and C.F. Nockels, 1973. The effect of vitamin E on egg production, hatchability and humoral immune response of chickens. Poult. Sci., 52: 778-783.
Direct Link - Yu, B.P., 1994. Cellular defenses against damage from reactive oxygen species. Physiol. Rev., 74: 139-162.
CrossRefDirect Link