
E. Rowghani
Department of Animal Science, College of Agriculture, Shiraz University, Shiraz, Iran
M. J. Zamiri
Department of Animal Science, College of Agriculture, Shiraz University, Shiraz, Iran
S. R. Ebrahimi
Department of Animal Science, College of Agriculture, Shiraz University, Shiraz, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 15 | Page No.: 2835-2840







ABSTRACT
Seventy Mehraban male lambs (initial live weight 43.9±4.3 kg) were used in a 70-day feeding experiment. Lambs were fed with 12 diets in a completely randomized design arranged in a 3x4 factorial trial with three levels of thiamin (0, 4 and 6 mg kg-1 DM) and 4 levels of monensin (0, 5, 11 and 22 mg kg-1 DM). A 21-day period was included for adaptation to the diets. Basal diet (dry matter basis) consisted of 7.5% corn silage, 8% alfalfa hay, 70% barley grain, 10% wheat bran, 3% cottonseed meal, 1.1% limestone and 0.4% vitamin and mineral supplement. Carcass characteristics, average daily gain and Feed Conversion Ratio (FCR) were not significantly (p>0.05) different between diets containing monensin and thiamin or their combinations with control. Daily dry matter intake was lowest (p<0.05) for diets 8 (11 mg monensin and 4 mg thiamin per kg DM) and 12 (22 mg monensin and 6 mg thiamin per kg DM) compared with diet 3 (6 mg thiamin per kg DM). BUN level and ruminal fluid pH were not significantly (p>0.05) different between diets. Blood glucose of lambs fed with 11 mg monensin/kg DM (50.6 mg dL-1) was higher (p<0.05) than control group (45.8 mg dL-1). Monensin tended to improve FCR. Monensin reduced DMI (p<0.05) and decreased feed consumption by 9.13-9.75% compared with the control diet. The effect of monensin on blood glucose concentration was significant (p<0.05) which was higher for two levels of monensin (11 and 22 mg kg-1 DM), compared with the control diet. Ruminal fluid pH was significantly (p<0.05) higher with all levels of monensin and at 22 mg kg-1 monensin, ruminal ammonia concentration was lowest (p<0.05). The overall effect of thiamin was a decrease (p<0.05) in BUN concentration.
PDF Abstract XML References Citation
How to cite this article
E. Rowghani, M. J. Zamiri and S. R. Ebrahimi, 2006. Effects of Monensin and Thiamin and their Combinations on Feedlot Performance, Blood Glucose, BUN Levels and Carcass Characteristics of Mehraban Lambs Fed a High Concentrate Diet. Pakistan Journal of Biological Sciences, 9: 2835-2840.
DOI: 10.3923/pjbs.2006.2835.2840
URL: https://scialert.net/abstract/?doi=pjbs.2006.2835.2840
DOI: 10.3923/pjbs.2006.2835.2840
URL: https://scialert.net/abstract/?doi=pjbs.2006.2835.2840
INTRODUCTION
Shortage of feed is the main constraint in livestock industry in Iran. High feed cost encourages the farmers to look for cheaper feeds or somehow reduce the feed conversion ratio. One method which may increase the feed efficiency is the addition of ionophores to the feed. Ionophores have been defined as substances capable of binding numerous mono-and divalent cations primarily in dimeric complexes that facilitate the passage of metal ions through hydrophobic lipid membrane (Ovehinnikov, 1979). Inophores vary in their affinity for metal ions.
Antibiotic ionophores such as monensin have been widely used to improve feed efficiency of cattle under both feedlot and grazing conditions (Oliver, 1975; Potter et al., 1976; Raun et al., 1976; Boling et al., 1977; Bergen and Bates, 1984) and in lambs (Glenn et al., 1977; Joyner et al., 1979) and in some cases, increased the daily gain (Calhoun et al., 1979). Monensin is a polyether ionophore produced by a strain of Streptomyces cinnamonensis that improves energetic efficiency of ruminal fermentation (Hyaney and Hoehn, 1967). Monensin is usually described as an antiporter that facilitates a one-for-one exchange of H+ and Na+ across cell membranes; however it can also mediate a K+ and H+ exchange (Pressman and Fahim, 1982). Bergen and Bates (1984) implied that monensin would lead to a depletion of interacellular Na+ rather than an increase, which in turn stimulates the ruminal environment.
Monensin alters ruminal fermentation in animals fed on starch-rich diets. Monensin acts on energy and nitrogen metabolism by decreasing molar proportion of acetic acid and increasing propionic acid (Richardson et al., 1976; Wedegaertner and Johnson, 1983; Jalc and Certik, 2005; Shinzato et al., 2006) and decreasing methane production (Fuller and Johnson, 1981; Guan et al., 2006) and also by increasing non-ammonia nitrogen flow in the duodenum (Russell and Strobel, 1988). It has been suggested that increased production of propionic acid would be energectically more efficient for the ruminant animal (Raun et al., 1976; Richardson et al., 1976), which is supported by the observed decrease in feed requirements for live weight gain that accompany the addition of monensin to feedlot diets (Raun et al., 1976; Boling et al., 1977). Jalc and Certik (2005) reported that the fermentation efficiency (E) and recovery of H2 were increased significantly by supplementing diet with monensin in artificial rumen. Also, monensin has inhibitory activity to gram-positive anaerobic bacteria like Streptococcus bovis (Muir and Barreto, 1979) that proliferates in the rumen when large amounts of grains are fed. Lactic acid production by S. bovis is responsible for the initiation of ruminal acidosis (Slyter, 1976). Acute deficiencies of thimain have been found in ruminants (Loew, 1975) and there are indications that subclinical deficiencies may occur (Brent, 1976). Brent (1976) stated that the development of lactic acidosis due to the feeding of rapidly fermentable materials is conductive to the initiation of polioencephalomalacia. Edwin et al. (1968) proposed that polioencephalomalacia was caused by a thiaminase enzyme. Davies et al. (1965) found the disease to be responsive to thiamin administration. Brethour (1972) found average daily gain of steers increased from 1.14 to 1.26 kg day-1 (p<0.05) due to the addition of 1 g thiamin and 100 g sodium bicarbonate to a wheat-based ration. Thiamin supplementation of an all-concentrate diet for feedlot steers resulted in higher feed intake and daily weight gain (Grigat and Mathison, 1982). The objectives of this study was to provide additional information on the possible benefit of the addition of monensin and thiamin to Mehraban male lambs (Iranian fat-tailed sheep) fed with a high concentrate diet.
MATERIALS AND METHODS
This experiment was carried out at the Animal Research Station, College of Agriculture, Shiraz University Iran. The experiment started on 25 July, 2005. Seventy Mehraban male lambs weighing 43.90±4.3 kg were randomly assigned to twelve groups of five and six lambs for two and ten diets respectively, in a 70 day feedlot experiment. Thiamin was fed at 0, 4 and 6 mg kg-1 of diet DM and monensin at 0, 5, 11 and 22 mg kg-1 of diet DM. A 21 day and 7 day adaption periods were included for the diets containing monensin and diets with no monensin, respectively. Lambs were fed twice daily and each diet was offered as a Total Mixed Ration (TMR). Water and a salt lick were available ad libitum. The animals were individually housed indoors in 1x1.5 m pens under normal lighting and their health was monitored continuously. Discarded feed was weighed and sampled for DM and subtracted from DM offered to calculate DMI. Ingredient and nutrient composition of the basal diet is presented in Table 1.
| Table 1: | Dietary ingredients and nutrient composition of the basal diet for lambs |
 | |
* Each kg contained: 500000 IU vitamin A, 100000 IU vitamin D3, 100 mg vitamin E, 190 g Ca, 90 g P, 19 g Mg, 50 g Na, 2 g Mn, 3 g Fe, 0.3 g Cu, 3 g Zn, 0.1 g Co and 1 mg Se | |
Lambs were weighed every three weeks after withdrawing food and water for 18 h and blood was collected from the jugular vein. The serum was separated after centrifugation at 5000 g for 15 min and stored at-20°C until analysed for blood glucose (using a commercial kit) and urea nitrogen (BUN) (Thomas, 1998). At the end of the experiment, lambs were slaughtered after withdrawing food and water for 15 h. Hot and cold (after 24 h) carcass weights were recorded. Subcutaneous fat depth was measured with a caliper at four points between the 12th and 13th ribs.
Rumen samples were collected using three 2-year old Mehraban rams fitted with ruminal fistula before feeding and again 0.5, 2 and 4 h after feeding. Feeding procedures and diets for fistulated rams were similar to those in the feedlot trial. Rumen fluid pH was measured directly in the whole rumen sample with a pH meter (Metrohem, Swiss). Rumen liquor samples were prepared by straining of the rumen sample through two layers of cheesecloth. The filtrate was treated with 5 mL of 0.2 mol L-1 HCl per 5 mL of filtrate and frozen at -20°C until measurement of NH3-N concentration (Kjeltec Auto Analyser 10304 Hogans, Tecator AB, Sweden).
Food and orts were analysed for DM and total nitrogen (N x6.25) according to the AOAC (1990) and Acid Detergent Fiber (ADF) and Neutral Detergent Fiber (NDF) were determined by procedures of Van Soest (1963) and Van Soest and Wine (1976), respectively.
Feedlot and carcass characteristics were analysed as a completely randomized design according to GLM procedures of SAS (1997) and serum glucose and urea nitrogen concentrations and ruminal pH and ammonia nitrogen were analysed according to Proc Mixed as Repeated Measures. Differences between the treatment means were tested using Tukey’s test.
RESULTS AND DISCUSSION
No death or digestive problems recorded. The effects of monensin and thiamin levels on the observed feedlot performance, blood parameters and ruminal fluid pH and ammonia concentration are presented in Table 2. Only daily Dry Matter Intake (DMI) was affected by treatments (p<0.05) among feedlot characteristics. Diets 8 (11 mg kg-1 monensin and 4 mg kg-1 thiamin) and 12 (22 mg kg-1 monesin and 6 mg kg-1 thiamin) were consumed significantly less than diet 3 (6 mg kg-1 thiamin), with no differences between other diets compared with diet 1. Lower DMI with feeding monensin was in agreement with other reports (Joyner et al., 1979). Lower DMI, at least in part, could be due to highter ruminal and blood propionic acid (Baile and Mayer, 1970), higher diet metabolizable energy due to lower loss of energy as methane (Wedegaertner and Johnson, 1983) and a longer retention time (decreased rumen turnover rate) of food in the rumen (Lemenager et al., 1978). Monensin did not show a dose related effect on feed intake, which is in agreement with the results of Stock et al. (1995) but not with the findings of Burrin et al. (1988), who reported decreases in feed intake as the monensin level increased.
Thiamin had no significant (p>0.05) effect on feed intake, in agreement with the findings of Grigat and Mathison (1982) who fed an all-concentrate diets supplemented with thiamin to feedlot steers. It seems that the higher ADF content of the diets (Mertens, 1997) and longer adaptation period (NRC, 2001), maintained higher ruminal pH and inhibited the activity of thiaminase producing bacteria (Brent, 1976). Average daily gain ranged 148-196 g, diet 1 had the lowest and diet 3 had the highest numerical values. Small differences in DMI and efficiency were observed between lambs fed 0, 5, 11 and 22 mg kg-1 monensin (Data not shown), which agrees with the all-concentrate, dry-rolled corn data of Stock et al. (1990). Although not significant, the highest and the lowest feed efficiencies were noted for diet 1 (11.40) and diet 8 (7.70), respectively. This was expected due to noticeable weight gain and less DMI with diet 8 (Table 2).
Improvement in feed efficiency has been reported by Grigat and Mathison (1982) with feedlot steers diet supplemented with 1.9 mg kg-1 thiamin which could be attributed to higher microbial protein synthesis (Hoeller et al., 1977). Feed efficiency values in the present study were higher than values reported by Joyner et al. (1979) with lambs fed with a 50% concentrate diet. The difference between the results might be due to stress during blood sampling, food withdrawing for 18h before periodical weighing and higher initial weight of lambs.
Blood glucose concentration was affected by treatments (p<0.05) and was higher for lambs diets supplemented with 11 and 22 mg kg-1 monensin compared with diet 1. BUN and ruminal fluid pH were not different among diets (p>0.05). Overall, thiamin significantly (p<0.05) decrease BUN levels as compared with the control diet (Data not shown). Improvement in ruminal fluid pH with feeding monensin (Table 3) could be a result of lower ruminal lactate production depressed growth of S. bovis (Dennis et al., 1981).
Monensin, irrespective of the thiamin levels resulted in a decrease in ruminal ammonia concentration (Table 2), which could be a result of inhibition of gram-positive bacteria by monensin and controlling the growth of ureolytic and proteolytic bacteria (Han et al., 2002).
Final weight was not different among treatments (p>0.05) in agreement with previous reports (Joyner et al., 1979; Stock et al., 1990). Although monensin can increase the net energy of the diet through increasing molar proportion of propionic acid (Sauer et al., 1989) and by reducing methane formation in the rumen (Okelly and Spiers, 1992), it also decreases feed intake which will not affect the final weight (Joyner et al., 1979; Stock et al., 1995). Joyner et al. (1979) fed lambs with diets containing 0, 10, 20 and 30 mg kg-1 monensin and reported 2-18% and 7-11% improvements in feed intake and feed efficiency, respectively, which might be due to higher molar proportion of propionic acid (Mass et al., 2001), lower methane production (Okelly and Spiers, 1992) and inhibition of protein degradation in the rumen (Poos et al., 1979).
Monensin did not significantly affect (p>0.05) the back fat thickness; however Fontenot and Huchette (1993) reported increased backfat thickness in cattle with monensin supplementation. This was not unexpected because higher fat production with feeding monensin could be related to the higher blood glucose and insulin concentration (Potter et al., 1976). The proportion of concentrate in this experiment was higher than that of Fontenot and Huchette (1993) trial and blood glucose was not increased dramatically with feeding monensin in present study. Blood glucose was significantly (p<0.05) higher with lambs fed monensin at 11 and 22 mg kg-1 compared with control diet. Blood gluocse ranged between 46.8-49 mg dL-1. The higher blood glucose with feeding monensin can be attributed to the higher molar percentage of propionic acid in the rumen (Benz et al., 1989). Non-dramatic changes in blood glucose could be due to the higher concentrate percentage in present study. In addition, the response to monensin supplementation of high concentrate diets is less than diets of high roughage in case of changes in molar proportion of propionic acid (Morris et al., 1990).
| Table 2: | Feedlot performance, blood glucose and urea nitrogen (BUN) concentrations (mg dL-1) and ruminal fluid pH and ammonia concentration (mg dL-1) of Mehraban lambs as influenced by the dietary levels of monensin and thiamin (mean±SE) |
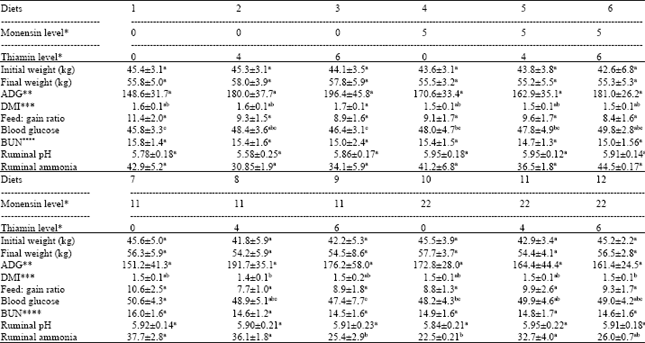 | |
a, b, c: Means in the same row with different superscript(s) are significantly different (p<0.05). * mg kg-1 diet DM. ** Average daily gain (g) *** Daily dry matter intake (kg/day) **** Blood urea nitrogen (mg dL-1) | |
| Table 3: | Blood glucose concentration (mg dL-1) and ruminal fluid pH and ammonia concentration (mg dL-1) of Mehraban lambs as influenced by the dietary levels of monensin (Mean±SE) |
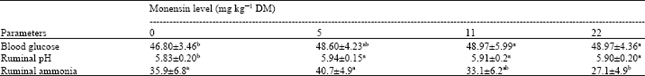 | |
| a, b Means in the same row with different superscript(s) are significantly different (p<0.05) | |
There was no significant effect of monensin on BUN concentration. Stephenson et al. (1997) reported that feeding monensin before calving had no any effect on BUN concentration but Poos et al. (1979) reported a decreasing ruminal ammonia and an increase with BUN in lambs fed monensin. Monensin at all levels (5, 11 and 22 mg kg-1) improved rumen pH (p<0.05) which could be due to the lowering lactate producing bacteria (Nagaraja et al., 1982). Ruminal fluid ammonia was lowest (p<0.05) with feeding 22 mg kg-1 monensin which could be a result of lowering the population of protozoa (Poos et al., 1979) and proteolytic bacteria (Russell and Strobel, 1988) in the rumen at this level.
Thiamin supplementation at all levels did not show any significant (p>0.05) effect on feedlot and carcass characteristics, which was not in agreement with findings of Brethour (1972) and Grigat and Mathison (1982). This difference might due to the higher ADF content of the diet (Mertens, 1997) and longer adaptation period (NRC, 2001) of this study. Also Lee et al. (1985) reported that rate and efficiency of gain by stressed beef calves were improved over a 28 day period by addition of a combination of vitamin E and a mixture of B-vitamins to a feedlot diet.
Thiamin had no effects (p>0.05) on blood glucose and ruminal pH. This was expected since different levels of thiamin had no effect on feed intake. Thiamin supplementation at both levels decreased (p<0.05) BUN concentration.
Based on the result and the cost of each kg live weight gain, according to local prices, supplementation of the diet 10-11 months old Mehraban male lambs with 11 mg monensin and 4 mg thiamin kg diet DM is more economical.
ACKNOWLEDGMENT
The authors would like to thank the financial support provided by Shiraz University Research Council, Project No. 83-AG-1703-C271.
REFERENCES
- Benz, D.A., F.M. Byers, G.T. Schelling, L.W. Greene, D.K. Lunts and S.B. Smith, 1989. Ionophores alter hepatic concentrations of intermediary carbohydrate metabolites in steers. J. Anim. Sci., 67: 2393-2399.
Direct Link - Bergen, W.C. and D.B. Bates, 1984. Ionophores: Their effects on production efficiency and mode of action. J. Anim. Sci., 58: 1465-1483.
Direct Link - Boling, J.A., N.W. Bradley and L.D. Campbell, 1977. Monensin levels for growing and finishing steers. J. Anim. Sci., 44: 867-871.
Direct Link - Brent, B.E., 1976. Relationship of acidosis to other feedlot aliments. J. Anim. Sci., 43: 930-935.
PubMed - Burrin, D.G., R.A. Stock and R.A. Britton, 1988. Monensin level during grain adaptation and finishing performance in cattle. J. Anim. Sci., 66: 513-521.
Direct Link - Calhoun, M.C., L.H. Carroll, C.W. Livingston and M. Shelton, 1979. Effect of dietary monesin on coccidial oocyst numbers, feedlot performance and carcass characteristics of lambs. J. Anim. Sci., 49: 10-19.
Direct Link - Dennis, S.M., T.G. Nagaraja and E.E. Bartley, 1981. Effects of lasalocid or monensin on lactate-producing or -using rumen bacteria. J. Anim. Sci., 52: 418-426.
PubMedDirect Link - Fontenot, J.P. and H.M. Huchette, 1993. Feeding sorbitol alone or in combination with monensin to finishing cattle. J. Anim. Sci., 71: 545-551.
Direct Link - Fuller, J.R. and D.E. Johnson, 1981. Monensin and lasalocid effects on fermentation in vitro. J. Anim. Sci., 53: 1574-1580.
CrossRefDirect Link - Guan, H., K.M. Wittenberg, K.H. Ominski and D.O. Krause, 2006. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci., 84: 1896-1906.
Direct Link - Han, H., H.S. Hussein, H.A. Glimp, D.H. Salyer and L.W. Green, 2002. Carbohydrate fermentation and nitrogen metabolism of a finishing beef diet by ruminal microbes in continous cultures as affected by ethoxyquin and (or) supplementation of monensin and tylosin. J. Anim. Sci., 80: 1117-1123.
Direct Link - Joyner, Jr., A.E., L.J. Brown, T.J. Fogg and R.T. Rossi, 1979. Effect of monensin on growth, feed efficiency and energy metabolism of lambs. J. Anim. Sci., 48: 1065-1069.
Direct Link - Lemenager, K.P., F.N. Owens, B.J. Shockey, K.S. Lusby and R. Totusek, 1978. Monensin effect on rumen turnover rate, twenty-four hour VFA pattern, nitrogen components and cellulose disappearance. J. Anim. Sci., 47: 255-260.
Direct Link - Mass, J.A., G.F. Wilson, S.N. Mc Cutcheon, G.A. Lynch, D.L. Burnham and J. France, 2001. The effect of season and monensin sodium on digestive characteristics of autumn and spring pasture fed to sheep. J. Anim. Sci., 79: 1052-1058.
Direct Link - Mertens, D.R., 1997. Creating system for meeting the fiber requirements of dairy cows. J. Dairy Sci., 80: 1463-1481.
Direct Link - Muir, L.A. and A. Barreto Jr., 1979. Sensitivity of streptococcus bovis to various antibiotics. J. Anim. Sci., 48: 468-473.
Direct Link - Nagaraja, T.G., T.B. Avery, E.E. Bartley, S.J. Galitzer and A.D. Dayton, 1982. Effect of lasalocid, monensin or thiopeptin on lactic acidosis in cattle. J. Anim. Sci., 54: 649-658.
CrossRefPubMedDirect Link - O'Kelly, J.C. and W.G. Spiers, 1992. Effect of monensin on methane and heat production of steers fed lucerne hay either ad libitum or at the rate of 250 g/h. Aust. J. Agric. Res., 43: 1789-1793.
CrossRefDirect Link - Oliver, W.M., 1975. Effect of monensin on gains of steers grazed on Coastal Bermudagrass. J. Anim. Sci., 41: 999-1001.
Direct Link - Poos, M.I., T.L. Hanson and T.J. Klopfenstein, 1979. Monensin effects on diet digestibility, ruminal protein bypass and microbial protein synthesis. J. Anim. Sci., 48: 1516-1524.
Direct Link - Potter, E.L., C.O. Cooley, L.F. Richardson, A.P. Raun and R.P. Rathmacher, 1976. Effect of monensin on performance of cattle fed forage. J. Anim. Sci., 43: 665-669.
Direct Link - Pressman, B.C. and M. Fahim, 1982. Pharmacology and toxicology of the monovalent carboxylic ionophores. Annu. Rev. Pharmacol. Toxicol., 22: 465-490.
PubMedDirect Link - Raun, A.P., C.O. Cooley, E.L. Potter, R.P. Rathmachez and L.F. Richardson, 1976. Effect of monensin on feed efficiency of feedlot cattle. J. Anim. Sci., 43: 670-677.
Direct Link - Richardson, L.F., A.P. Raun, E.L. Potter, C.O. Cooley and R.P. Rathmacher, 1976. Effect of monesin on rumen fermentation in vitro and in vivo. J. Anim. Sci., 43: 657-664.
Direct Link - Russell, J.B. and H.J. Strobel, 1988. Effects of additives on in vitro ruminal fermentation: A comparison of monensin and bacitracin, another gram-positive antibiotic. J. Anim. Sci., 66: 552-558.
Direct Link - Sauer, F.D., J.K.G. Kramer and W.J. Cantwell, 1989. Antiketogenic effects of monensin in early lactation. J. Dairy Sci., 72: 436-442.
CrossRef - Stephenson, K.A., I.J. Lean, M.L. Hyde, M.A. Curtis, J.K. Garvin and L.B. Lowe, 1997. Effects of monensin on the metabolism of periparturient dairy cows. J. Dairy Sci., 80: 830-837.
Direct Link - Stock, R.A., M.H. Sindt, J.C. Parrott and F.K. Goedeken, 1990. Effects of grain type, roughage level and monensin level on finishing cattle performance. J. Anim. Sci., 68: 3441-3446.
Direct Link - Stock, R.A., S.B. Laudert, W.W. Stroup, E.M. Larson, J.C. Parrott and R.A. Britton, 1995. Effect of monensin and monensin and tylosin combination on feed intake variation of feedlot steers. J. Anim. Sci., 73: 39-44.
Direct Link - Wedegaertner, T.C. and D.E. Johnson, 1983. Monensin effects on digestibility, methanogenesis and heat increment of cracked corn-silage diet fed to steers. J. Anim. Sci., 57: 168-177.
Direct Link - NRC, 2001. Nutrient Requirements of Dairy Cattle. 7th Edn., National Academy Press, Washington, DC, ISBN: 978-0-309-06997-7, Pages: 405.
CrossRefDirect Link - Shinzato, I., K. Abe, S. Kogawa, Y. Toride and J.T. Takahashi, 2006. Effects of L-cysteine supplementation on growth performance and ruminal fermentation in growing steers in comparison to monensin. Proceedings of the 2nd International Conference on Greenhouse Gases and Animal Agriculture, September 20-24, 2006, Zurich, Switzerland, pp: 189-192.
CrossRef - Morris, F.E., M.E. Branine, M.L. Galyean, M.E. Hubbert, A.S. Freeman and G.P. Lofgreen, 1990. Effect of rotating monensin plus tylosin and lasalocid on performance, ruminal fermentation and site and extent of digestion in feedlot cattle. J. Anim. Sci., 68: 3069-3078.
CrossRefPubMedDirect Link