
Sh. Enteshari
Department of Biology, University for Teacher Education, Tehran, Iran
Kh. Kalantari
Department of Biology, Shahid Bahonar University, Kerman, Iran
M. Ghorbanli
International Center of Science, High Technology and Environmental Sciences, Kerman, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 2 | Page No.: 231-237







ABSTRACT
The effect of different bands of UV-radiation and epibrassinosteroid treatments on the quantity of chlorophyll a,b, total chlorophyll, anthocyanine, carotenoid, flavonoids and UV absorbing compounds in leaves of the Glycine max were studied. In this study plants were sprayed with epibrassinosteroid and treated with UV-A (2.73 Wm-2), UV-B (1.15 Wm-2) and UV-C (2.66 Wm-2). Studied showed that in those plants which were treated with UV-A, the contents of these pigments were not significantly different when compared with control. While irradiation of the plants with UV-B and UV-C significantly caused decrease in chlorophyll a, b, total chlorophyll and carotenoids. Epibs treatment moderated chlorophyll and carotenoid reduction in those plants that treated with UV-B and UV-C. Decrease in carotenoid were less significant when compared with chlorophyll. The quantity of antocyanines, flavonoids and UV absorbing compounds in plants that treated with UV-B,UV-C and epibs were significantly increased. These pigments have high absorption in UV spectrum of light and therefore could be used as UV-absorbing compounds by plants to prevent penetration of UV to the more sensitive tissues. The role of flavonoids in quenching of Hydrogen peroxide and other species of active oxygen could well describe. The defensive reaction of plants against UVradiation and epibrassinosteroid will discussed in this study.
PDF Abstract XML References Citation
How to cite this article
Sh. Enteshari, Kh. Kalantari and M. Ghorbanli, 2006. The Effect of Epibrassinosteroid and Different Bands of Ultraviolet Radiation on the Pigments Content in Glycine max L. Pakistan Journal of Biological Sciences, 9: 231-237.
DOI: 10.3923/pjbs.2006.231.237
URL: https://scialert.net/abstract/?doi=pjbs.2006.231.237
DOI: 10.3923/pjbs.2006.231.237
URL: https://scialert.net/abstract/?doi=pjbs.2006.231.237
INTRODUCTION
The anthropogenic and natural destruction of stratospheric ozone has been linked to the increment in ultraviolet radiation at the earth’s surface (Hofmann et al., 2000). Plants like other livings, are deleteriously affected by UV-radiation. Photosynthetic processes, reduction of the photosynthetic pigments, degradation of proteins and DNA and increase oxidative stress are the main damage caused by UV radiation (Allen et al., 1998).
Sensitivity to UV-radiation is depend on plant species and cultivars, developmental stage and experimental conditions (Al-Oudat et al., 1998). Protective responses are also stimulated by UV-radiation including increase production of UV-absorbing compounds and secondary compounds including hydroxy cinnamic acid derivatives, phenylpropanoids and flavonoids which effectively will absorb the UV spectral (Hofmann et al., 2000; Middleton; Termura, 1993). Flavonoids are frequently found in or on epidermal layers where they have been shown to increase markedly following UV treatment. Studies with flavonoid mutants which are not able to synthesis flavonoid further high lighted the importance of flavonoids for UV-tolerance (Li et al., 1993).
Anthocyanins also have a wide distribution in mature and senescent leaves. These compounds generally have two maximum absorption, one between 240-290 nm and the other in the visible spectrum at 500-550 nm. The strong UV-absorption of anthocyanins has led to the hypothesis that these compounds may protect leaves from the harmful effects of UV-radiation (Woodall and Stewart, 1998).
Brassinosteroids are unique steroidal phytohormones in plant. They exhibit abroad spectrum of physiological functions (Bajguz, 2000a). Stem elongation, leaf development, xylem differentiation, pollen tube growth and senescence are influenced by these compounds (Tanaka et al., 2003). On the other hand studies shown that these compounds have an anti-stress effect on different plants (Bajguz, 2000b). For instance it was shown that brassinosteriod help to overcome stress caused by low and high temperatures (Khripach et al., 2000), drought (Salram, 1994) heavy metals (Bajguz, 2000a), infection (Li and Chory, 1999) and salt stress (Ozdemir et al., 2004).
In this research we studied the effect of different bands of UV radiation on photosynthetic pigments, anthocyanins, flavonoids and UV-absorbing compounds in soybean leaf and elucidate wether or not the increased UV-radiation resistance by brassinosteriod is mediated via the oxidative system in leaves of Glycine max (cultivar williams) when subjected to UV- radiation. For this purpose we examine the effect of 24-epibrassinolide (24-epibs) and different bands of UV radiation on photosynthetic pigments, flavonoids, anthocyanins and UV- absorbing compounds in soybean leaf.
MATERIALS AND METHODS
Plant growth: Soybean seeds (Glycine max L. cv williams) were obtained from Gorgan Agricultural Research Institute-Iran, sterilized for 20 min with 5% (w/v) sodium hypochlorite solution and then thoroughly rinsed with deionized water. They were sown in 15x15 cm Pots containing vermiculite and irrigated routinely with Hogland’s solutions. Plants were grown in a growth chamber, (Grouc-company) during the 16 h day and 8h night cycle. Temperatures were 27±2°C during the day and 23±2°C at night. The intensity of light above the surface of plant was 11000 lux.
Approximately 16 days after sowing seeds, when the secondary trifoliate blades of plants had emerged were used for treatments.
For UV-A UV-B and UV-C treatment two UV-A (philips, 18 w λ = 380 nm) lamps, two UV-B (philips, 15 w λ = 312 nm) lamps and one UV-C (philips, 30 w λ = 254) lamp were used. Plants were exposed to UV-A and UV-B for 20 min every day lasting 8 days. For UV-C treatment, plants were exposed to this radiation 3 min for 4 days. Intensity of light just above the plant, for UV-A, UV-B and UV-C treatments were 2/45 w/m–2, 1/15 w/m–2 and 2/6 w/m–2, respectively.
Brassinosteriod treatment: Epibrassinolid (sigma E- 1641) at 10–7 M containing 0/01% tween 20 (polyoxy ethylene sorbitan) was used to spray. Plants were sprayed three days before UV treatment. The control plants were sprayed with distilled water and tween 20.
Chlorophyll and carotenoid quantification: Chlorophyll pigments were extracted in 80% acetone in the dark for 24 h at 4°C. The extracts were centrifuged and absorbances measured with a UV/visible spectrophotometer (2100-WPA) at 470, 648, 663 nm and then equations described by Lichtenthaler (1987). Were used for quantification of these compounds.
Anthocyanin quantification: Anthocyanin concentration measured by Wagner method (1979).
Flavonoids and UV- absorbing compounds: To determine the absorption of flavonoids and other UV-absorbing compounds, fresh leaf discs were extracted in ethylalchol: acetic acid (99:1,V:V). Then boiled for 10 min in a water bath at 80°C and centrifugated. The absorbance was read at 300 nm with UV/visible spectrophotometer (2100WPA). For flavonoid determination dried leaf discs were extracted in ethyl alchol:acetic acid (99:1, V:V). The absorbance was read at three wave lengths: 270, 300 and 330 nm (Krizek et al., 1998).
Statistical analyses: All analyses were done on a completely randomized design. Data were analysed by the Analysis of Variances followed by Duncan Test. Each data was the mean of six replicates and means were considered as significant differences at p≤5% level. All statistical analyses were performed using the SAS for Windows.
RESULTS
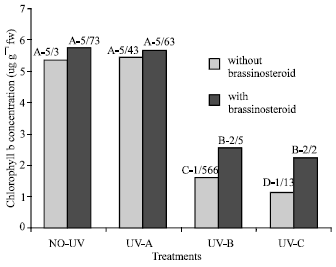
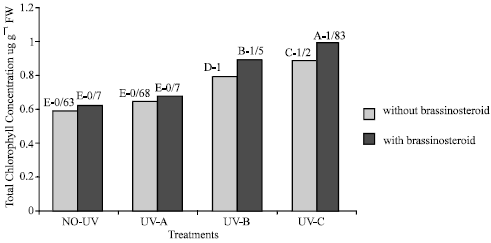
Chlorophyll: Chlorophyll content (a, b and total) of plants which were exposed to UV-B and UV-C decreased significantly (Fig. 1-3), but chlorophyll content of those plants which were exposed to UV-A did not change significantly (p<5%). Epibs treatment moderated chlorophyll reduction in those plants that treated with UV-B and UV-C (p<5%).
Carotenoid: UV-B and UV-C caused significant decrease in the concentration of carotenoid (p<5%).
 | |
| Fig. 1: | Effect of different bands of UV radiation and epibrassinosteroid treatments on total chlorophyll (a+b) content (ug g–1 FW) of soybean (p<5%) |
 | |
| Fig. 2: | Effect of different bands of UV radiation and epibrassinosteroid treatments on chlorophyll b content (ug g–1FW) of soybean (p<5%) |
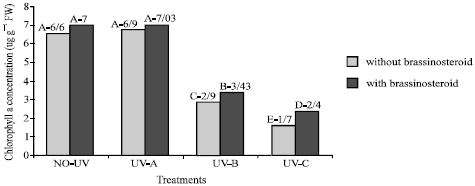 | |
| Fig. 3: | Effect of different bands of UV radiation and epibrassinosteroid treatments on chlorophyll a content (ug g–1 FW) of soybean (p<5%) |
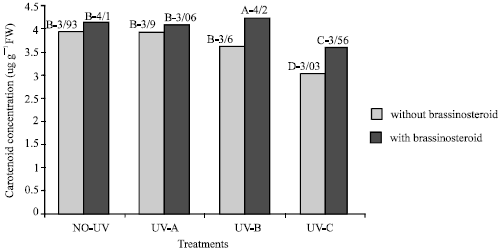 | |
| Fig. 4: | Effect of different bands of UV radiation and epibrassinosteroid treatments on carotenoid content (ug g–1 FW) of soybean (p<5%) |
Plants that treated with epibs moderated this decrease (Fig. 4). UV-A did not have any significant effect on carotenoid content (p<5%) (Fig. 4).
Flavonoid and UV-absorbing compound: Extracts of fresh and dry leaves taken from plants that were treated with UV-B and UV-C radiation showed significant increase in UV absorbance percentage at 270, 300 and 330 nm (Fig. 5-8). Application of epibs significant increased UV-absorbance percentage at these wave-lengths in plants that treated with UV-B and UV-C (p<5%) (Fig. 5 to 8) but UV absorbing percentage of those plants which were exposed to UV-A did not change significantly.
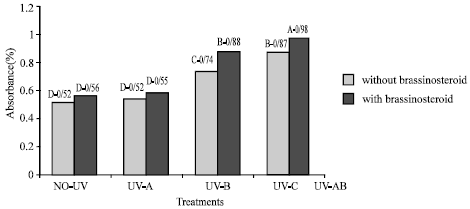 | |
| Fig. 5: | Effect of different bands of UV radiation and epibrassinosteroid on UV absorbing compound absorbance of soybean (p<5%) |
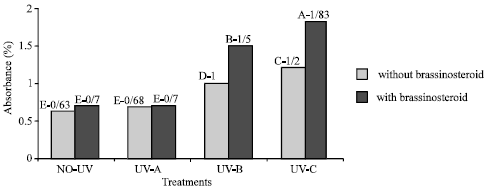 | |
| Fig. 6: | Effect of different bands of UV radiation and epibrassinosteroid treatment on flavonoid absorbance at 330 nm of soybean (p<5%) |
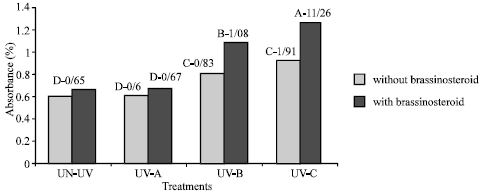 | |
| Fig. 7: | Effect of different bands of UV radiation and epibrassinosteroid treatment on flavonoid absorbance at 300 nm of soybean (p<5%) |
Anthocyanins: Those plants which were exposed to UV-B and UV-C radiation showed significant increase in anthocyanins contents of leaves. But anthocyanins content of these plants which were grown in presence of UV-A radiation did not increase. Epibs application increased anthocyanins in those plants which were exposed to UV-C and UV-B radiation (Fig. 9) (p<5%).
DISCUSSION
The concentration of chlorophyll a, chlorophyll b, total chlorophyll (a+b) and carotenoid were decreased when soybean plants were exposed to UV-B and UV-C radiation (Fig. 1-3). This result is in accordance with a report of Takeuchi et al., (2002). They reported that supplementary UV-radiation caused reduction in photosynthetic pigments in rice. However Barsig and Malz (2000) reported that in sugar maize leaves under enhanced UV-B radiation a slight decrease in chlorophyll b was observed but there was no significant differences on chlorophyll or carotenoid contents.
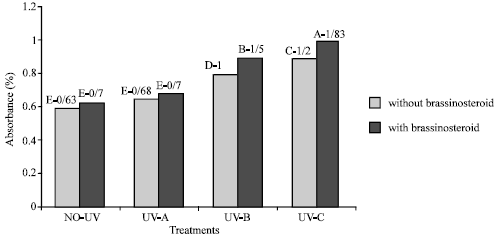 | |
| Fig. 8: | Effect of different bands of UV radiation and epibrassinosteroid treatment on flavonoid absorbance at 270 nm of soybean (p<5%) |
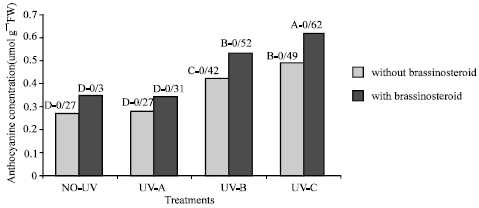 | |
| Fig. 9: | Effect of different bands of UV radiation and epibrassinosteroid treatments on anthocyanine content (umol g–1 FW) of soybean (p<5%) |
On the other hand, Yanqun et al. (2003) reported that responses of photosynthetic pigments of 20 soybean cultivars under UV-B radiation were complex because chlorophyll a content in some of these cultivars strongly decreased and chlorophyll b content less effected, but in numerous of these cultivars an increase in chlorophyll a/b ratio reported. It has been reported that supplementary UV-B radiation caused reduction in the chloroplast proteins, especially ribulose-1, 5 bis phosphate carboxylas oxygenase (rubisco) and light harvesting chlorophyll a/b binding protein of PS II (Mackerness et al., 1997) Caldwell et al. (1995) concluded that in most of the sensitive plants UV-B and UV-C radiation significantly decreased chlorophyll contents primarily because UV-B radiation destroyed the structure of chloroplast, inhibits synthesis of chlorophyll and destroyed the structure of the membrane of chloroplast and increased the rate of degradation of chlorophyll. The efficiency of carotenoids in protecting the photosystems is likely due to their function as efficient quenchers of high energy of short wave radiation (Middleton and Termura, 1993). Mechanism by which this is accomplished was first proposed to involve a photochemical state change from singlet oxygen to triplet form by interaction with carotenoids, removing the potentially dangerous oxygen radicals produced in photo oxidative processes (Inze and Montagu, 2002). In this study application of epibs increased the chlorophyll and carotenoid content either in those plants which were treated with UV-A radiation or plants which were not (Fig. 1-3). In those plants that were treated with epibs and exposed to UV-B and UV-C, carotenoid and chlorophyll decrease was moderated. Studies have shown that carotenoids serve a protective function against UV-B and UV-C radiation (Middleton and Termura, 1993). In the other hand Liydmila (2002) reported that application of epibs to the Arabidopsis plant which pretreated with cadmium, repairment of the chlorophyll loss was significant. Khripach et al. (1999) concluded that epibs possibly act through phytochrome reactivation has capacity to improve chlorophyll loss that caused by cadmium toxicity. They reported that a short treatment of triticale seedling with red or with a combination red and far-red light increased the chlorophyll and protochlorophyll content significantly. However during dark incubation the content of pigments increased till about 28 h of incubation and after that the total pigment decreased. When the effects of successive irradiation by red and far red light and those of only a red-light irradiation on pigment synthesis were compared, plants that treated with epibs and untreated reacted differently. This result probably means that far-red light effect on phytochrom and photosynthetic pigments was counter balanced partially by the action of epibs (Khripach et al., 1999).
In this study we observed that the concentration of anthocyanins in leaf that exposed to UV-radiation increased. This results are similar to those reported for Syzgium by Woodall et al., (1998). They reported high anthocyanin concentrations in mesophyll of juvenile Syzygium leaves which may indicated that anthocyanines are most important UV-absorbing compounds especially if the cuticle and epidermis do not fully attenuate incident of UV-radiation. In the other hand Takahashi et al. (1991) reported that anthocyanins protect cell suspension cultures of Centaurea cyanus from the damaging effect of UV-radiation. The induction of anthocyanins by environmental stresses have prompted many workers to postulate roles for anthocyanins in leaves. The various functional hypotheses that have been debated in recent years are (I) modification of the quantity and quality of captured light (II) protection from the effects of UV-B (III) defence from herbivory (IV) protection from photo inhibition (V) scavenging of reactive oxygen intermediates under stressful environments (Gould et al., 2000).
Flavonoids and UV-absorbing compounds can increase rapidly in response to UV-B and UV-C radiation. The involvement of flavonoids in the UV-B response has been reported for several plant species including Trifolium repense (Hofmann et al., 2000) New red fire lettuce (Krizek et al., 1998), Glycine max (Yanqun et al., 2003) and so on. Accumulation of flavonoids is frequently observed in response to UV-B radiation and has been linked to UV-B absorption characteristic displayed by flavonoids and UV-absorbing compound as radical scavenging antioxidants (Hofmann et al., 2000). Gitz et al. (2004) shown in soybean cultivars which treated with UV-B the maximal absorption consistent with hydroxycinnamic acids compound. They concluded that this compound was serving as UV-absorbing compounds. In the other hand Sharma et al. (1998) reported that in wheat seedings which were exposed to UV-B flavonol (kampherol, caumarin), flavone, anthocyanin and cinamic acid increased.
In this study we observed that epibs increased anthocyanins, flavonoids and UV-absorbing compounds in soybean plants that treated with UV-B and UV-C treatment Krishna (2003) reported that epibs enhanced plant resistance against fungal pathogen infection. The increase resistance was associated with enhancement of abscisic acid, ethylene levels and the presence of phenolic and terpenoid substances. In this study they reported that increment in activity of peroxidase and polyphenoloxidase enzymes involved in the metabolism of poly phenols in Cucumber. The application of epibs can protect plants against UV-radiation by increasing protective pigments such as flavonoids and anthocyanins which could be generated by polyphenols synthesis pathway. Li et al., (1993) showed that Arabidopsis thaliana mutants which are defective in ability to synthesis UV-B absorbing compounds (flavonoids and sinnapic esters) were more sensitive to UV-B than wild type, indicating a role for flavonoids in protecting plants from UV-B damage. Sharma et al. (1998) reported that anthocyanins not only chelates metal ions but also forms ascorbic acid (co-pigment)-metal-anthocyanin complex which could scavenge hydro-peroxy radicals. Hallbrok et al. (1976) have shown an increased expression of genes that encoding enzymes such as phenylalanine amonialyase (PAL), chalcone synthetase (CHS) of the phenyl propanoid and flavonoid biosynthesis. Therefore we guessed that epibs effected these enzymes increased flavonoid, UV-absorbing and anthocyanin synthesis.
In conclusion from the difference in alteration of photosynthetic pigments, flavonoids, anthocyanins and UV-absorbing compounds in soybean may suggested that epibs increased plant resistance against oxidative stress and UV radiation. However, for definitive conclusion, a comparison between UV-sensitive and UV-resistant varieties in terms of induction of protective pigments, photosynthetic pigments and antioxidant enzymes activities might be enlightening for anti-stress role of epibs.
REFERENCES
- Bajguz, A., 2000. Blockade of heavy metals accumulation in Chlorella vulgaris cells by 24-epibrassinolide. Plant Physiol. Biochem., 38: 797-801.
CrossRefDirect Link - Barsig, M. and R. Malz., 2000. Fine structure carbohydrates and photosynthetic pigments of sugar maize leaves under UV-B radiation. Environ. Exp. Bot., 43: 121-130.
Direct Link - Caldwell, M.M., A.H. Teramura and M. Tevini, 1995. Effects of increased solar ultraviolet radiation on terrestrial plants. Ambio, 14: 166-173.
Direct Link - Gould, K.S., K.R. Markham, R.H. Smith and J.J. Goris, 2000. Functional role of anthocyanins in the leaves of Quintinia serrata A. Cunn. J. Exp. Bot., 51: 1107-1115.
CrossRefPubMedDirect Link - Gitz, D.C., L.L. Gitz, S.J. Britz and J.H. Sullivan, 2004. Ultraviolet-B effects on stomatal density, water use efficiency and stable carbon isotope discrimination of four glasshouse grown soybean (Glycine max) cultivars. Environ. Exp. Bot., 53: 343-355.
Direct Link - Hallbrok, K., K.H. khobloch, F.J. Kreuzaler, R.M. potts, and E. wellmann, 1976. Coordinated induction and subsequent activity changes of two groups of metabolically interrelated enzymes light induced synthesis of flavonoid glycosides in cell suspension cultures of petroselinume horetense. Eur. J. Biochem., 61: 199-200.
- Hofmann, W., E.E. Swing, S.J. Bloork, R.R. Markham, K.G. Ryan, B.D. Campbell, B.R. Jordan and D.W. Fountain, 2000. Responses of nine Trifolium repens L. Populations to ultraviolet-B radiation: Differential flavonol glycoside accumulation and biomass production. Ann. Bot., 86: 527-573.
Direct Link - Khripach, V., V. Zhabinskii and A. de Groot, 2000. Twenty years of brassinosteroids: Steroidal plant hormones warrant better crops for the XXI century. Ann. Bot., 86: 441-447.
CrossRefDirect Link - Krishna, P., 2003. Brassinosteroid-mediated stress responses. J. Plant Growth Regul., 22: 289-297.
CrossRefDirect Link - Krizek, D.T., S.J. Britz and R.M. Mirecki, 1998. Inhibitory effects of ambient levels of solar UV-A and UV-B radiation on growth of cv. new red fire lettuce. Physiol. Plant., 103: 1-7.
CrossRefDirect Link - Middleton, E.M. and A.H. Termura, 1993. The role of flavonol glaycosides and carotenoids in protecting soybean from ultraviolet-B damage. Plant Physiol., 103: 741-752.
Direct Link - Ozdemir, F., B. Melike, T. Demiral and I. Turkan, 2004. Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress. Plant Growth Regul., 42: 203-211.
CrossRefDirect Link - Salram, R.K., 1994. Effects of homobrassinolide application on plant metabolism and grain yield under irrigated and moisture-stress conditions of two wheat varieties. Plant Growth Regul., 14: 173-181.
CrossRefDirect Link - Sarma, D., Y. Sreelakhsmi and R. Sharma, 1997. Antioxidant ability of anthocyanins against ascorbic acid oxidation. Phytochemestry, 45: 671-674.
CrossRef - Takeuchi, A., T. Yamaguchhi, J. Hidema, A. Stride and T. kumagai, 2002. Changes in synthesis and degradation of rubisco and LHC II with leaf age in rice (Oryza sativa L.) growing under supplementary UV-B radiation. Plant Cell Environ., 25: 695-706.
Direct Link - Tanaka, K., Y. Nakomura, T. Asami, Sh. Yoshida, T. Matsuo and S. Okamoto, 2003. Physiological roles of brassinosteroids in early growth of Arabidopsis: Brassinosteroids have a synergistic relationship with gibberellin as well as relationship with gibberellin as well as auxin in light-grown hypocotyls elongation. J. Plant Growth Regul., 32: 259-271.
Direct Link - Wagner, G.J., 1979. Content and vacuole/extravacuole distribution of neutral sugars, free amino acids and anthocyanin in protoplasts. Plant Physiol., 64: 88-93.
PubMed - Yanqun, Z., L. Yuan, C.J. Haiyan and C. Lanjun, 2003. Intraspecific differences in physiological response of 20 soybean cultivars to enhanced ultraviolet-B radiation under field conditions. Environ. Exp. Bot., 50: 87-97.
Direct Link