
M. Mahzounieh
Department of Pathobiology, Faculty of Veterinary Medicine, Shahrekord University, Iran
E. Moghtadaei
Graduates From Faculty of Veterinary Medicine, Shahrekord University, Iran
T. Zahraei Salehi
Department of Microbiology and Immunology, Faculty of Veterinary Medicine,
Tehran University, Iran
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 2 | Page No.: 227-230







ABSTRACT
Caliciviruses may cause a wide spectrum of disease in animals and are important etiological agent of viral gastroenteritis in humans. Members of the family caliciviridae are small nonenveloped viruses 27 to 35 nm in diameter. They possess a single stranded poly adenylated RNA genome. Caliciviruses have been isolated from mink, dog, cattle and non-human primates. Norwalk-like Viruses (NLVs) are the most common cause of acute non-bacterial gastroenteritis in humans. Cattle may be a reservoir of NLVs although never bovine NLVs have been found in humans. In this study, we try to detect enteric caliciviruses genome from 50 fecal samples of 6 dairy cattle herds in Shahrekord area using reverse transcriptase polymerase chain reaction (RT-PCR) assays specific for NLVs found in humans. The primers used for PCR amplification were Ni and E3, which amplify a 113 bp product for the detection of both genogroups I and II SRSV RNA in fecal material. Our results showed that nine specimens (18%) were positive. These findings suggest that calicivirus infection is endemic in dairy herds in Shahrekord, Iran and may be have an important role in calf diarrhoea.
PDF Abstract XML References Citation
How to cite this article
M. Mahzounieh, E. Moghtadaei and T. Zahraei Salehi, 2006. Detection of Calicivirus Genome in Calves Using Ni/E3 Primers in Shahrekord Area, Iran. Pakistan Journal of Biological Sciences, 9: 227-230.
DOI: 10.3923/pjbs.2006.227.230
URL: https://scialert.net/abstract/?doi=pjbs.2006.227.230
DOI: 10.3923/pjbs.2006.227.230
URL: https://scialert.net/abstract/?doi=pjbs.2006.227.230
INTRODUCTION
Caliciviruses (family Caliciviridae) are nonenveloped viruses, 27 to 35 nm in diameter with single-stranded RNA positive-strand genomes of 7 to 8 kb (Smiley et al., 2003). Electron microscopy has revealed that several, but not all species, display typical cup-shaped surface depression, the characteristic calyx morphology, from which the family derives its name (Prasad et al., 1999). Many investigators have reported the isolation of caliciviruses from diverse species. They are a well-established cause of respiratory, vesicular and hemorrhagic diseases in animals. In addition, these viruses are an important cause of enteric diseases in humans. Recently, molecular analysis of several bovine enteric calicivirus isolates indicated that they are genetically close to human enteric calicivirus. Enteropathogenic Bovine Enteric Caliciviruses (BECs) with small round-structured virus morphology, which are similar to human NLV, have also been identified in cattle. In recent years, two such NLV-BEC strains from Europe, Jena (Liu et al., 1999) and Newbury Agent-2 (NA-2) (Dastjerdi et al., 2002), were genetically characterized and shown to be similar to human NLVs. Similar virus gene sequences were also detected in pooled fecal samples collected from veal calves in The Netherlands (Van der Poel et al., 2000). These findings have raised questions about the host range of BECs because it was unclear whether these viruses comprised their own distinct genetic lineage or whether they were, potentially, part of a common pool of viruses circulating between animals and humans (Dastjerdi et al., 1999, 2002). The Norovirus genus of the Caliciviridae encompasses viruses that cause outbreaks of gastroenteritis in human and viruses that have been associated with diarrhoea in cattle. The two bovine noroviruses, Bo/Newbury2/76/UK and Bo/Jena/80/DE, represent two distinct genetic clusters in the newly described genogroup III (Oliver et al., 2003, 2004). To investigate if bovine enteric caliciviruses circulate in Shahrekord, Iran, this study was carried out between 2004 and 2005.
MATERIALS AND METHODS
Fecal preparation: Fecal samples were collected from affected calves and 10% suspensions of feces were prepared in 2 mL of DNase and RNase free double distilled water (ddH2O).
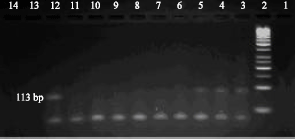 | |
| Fig. 1: | Lane 1- Negative control, 2- size marker, 3- Positive control, 4, 5 and 12 positive samples, 6, 7, 8, 9, 10, 11, 13 and 14 negative samples. |
The remaining material was stored at -20 until it tested by RT-PCR. Samples were collected from calves that age were 1-2 weeks to 2 month. The samples were collected from 30 male and 20 female calves.
Oligonucleotide primers: The primers used for PCR amplification were Ni (5’-GAA TTC CAT CGC CCA CTG GCT-3’) and E3 (5’-ATC TCA TCA TCA CCA TA-3’), which amplify a 113-bp product for the detection of both genogroup I and II SRSV RNA in fecal material (Green et al., 1995; Maguire et al., 1999).
RNA extraction and RT-PCR: RNA was extracted from 100 μL of the 10% fecal suspension by a method modified from that of Boom et al. (1990) with guanidine isothiocyanate and silica and eluted in 26 μL of RNase free water containing DEPC as a RNase inhibitor. One micro liter of primer was added to the extracted RNA, the mixture was heated at 70°C for 5 min and chilled on ice and 14 μL of the mixture was added. yielding a total volume of 35 μL of the PCR mixture consisting of 20 mM tris- HCl (pH. 8.4), 50 Mm KCl, 5 Mm MgCl2, each deoxy nucleotide triphosphate (dNTPs; Fermentas) at a concentration of 50 micromolar and 200 μ of RT. Reverse transcription was performed at room temperature for 10 min followed by incubation 37°C for 1 h, after which the reaction was terminated by incubation at 95° for 5 min followed by chilling on ice. Five microliter of cDNA was then added to 45 μL of the PCR mixture, yielding a total of 50 mL of a reaction mixture consisting of 18 mM Tris-HCl (pH. 8.4), 45 Mm KCl, 2 Mm MgCl2, each dNTPs at a concentration of 50 μM and 200 U of moloney murine leukemia virus (M-MuLV) RT. A DNA Thermal Cycler (Corbett Research) was programmed for 95°C for 15 min and 40 cycles of 94°C for 15 sec, 40°C for 45 sec and 72°C for 1 min, with a final extension at 72°C for 10 min. PCR amplicons were resolved on 2.5% agarose gels. A positive signal for primer pair NI/E3 was a 114 bp amplicon (Maguire et al., 1999).
Amplification products were examined by gel electrophoresis of 10 μL of reaction mixture in agarose gels containing ethidium bromide (4 μg mL–1 of agarose gel; Fig. 1). Appropriate positive and negative controls to confirm the sensitivity and specificity of the PCR and to detect contamination were included in each assay run. Nuclease-free water was used as a negative control for every five fecal samples in both the reverse transcription (RT) and PCR steps and was handled simultaneously.
RESULTS
During the epidemic season, we collected 50 fecal samples from six dairy cattle herds in Shahrekord area. The ages of calves were between 2 weeks to 2 month. These samples collected randomly and stored at -20°C until it was tested by RT-PCR. RT-PCR screening was performed with the broadly reactive primer pair Ni-E3. After amplification of extracted RNA genome by RT-PCR, nine samples out of 50 specimens were positive for the NiE3 primers. Therefore, prevalence of calicivirus infection was 18% in these herds. Six samples out of 30 male (5%) and 3 out of 20 female calves (6.6%) were found positive. There was no significant difference between male and female infection rate (p>0.05). The average age of positive calves was 17 days.
DISCUSSION
Caliciviruses are emerging pathogens that are a leading cause of acute epidemic gastroenteritis (Green et al., 1997; Mead et al., 1999). Human caliciviruses (HuCVs) are currently estimated to be responsible for >90% of food- and water-borne viral gastroenteritis in humans of all ages (Mead et al., 1999). In animals, caliciviruses cause a spectrum of diseases that include gastroenteritis (pigs, calves, dogs and chickens), vesicular lesions and reproductive failure (pigs and sea lions), rhinitis and conjunctivitis (cats and cattle), a fatal hemorrhagic disease (rabbits) and a stunting syndrome (chickens) (Thiel et al., 1999). Recently, RT-PCR with broadly reacting primers has been introduced. Information is lacking on the BEC infection patterns in cattle farms in Iran.
The Ni-E3 primer pair detects approximately 90% of SRSV genotypes circulating in the United Kingdom (Green et al., 1995), with increased sensitivity compared with that of EM. SRSV RNA could be detected in up to 30% of EM-negative specimens from outbreaks (Maguire et al., 1999) Also the present Ni/E3 PCR assays could be used to screen NLV-BECS. In this study, we used these primers to detect them in fecal calf samples in Iran. Our results showed that nine of 50 samples (18%) were positive in dairy herds. Smiley et al. (2003) used P289/290 primers in England. In two farms A and B, 14.5 % of samples were positive and genetic relationship between NLV-BEC and NLV- HuCVs were supported.
Two genetically distinct bovine enteric caliciviruses (BECs) have been identified using six RT-PCR primer sets designed for the detection of human NLVs or BECs by Smiley et al. (2003) the norovirus (NLV) Jena and Newbury Agent-2 (NA-2) BECs, which are genetically related to human noroviruses and the Nebraska (NB) BECs, which are related to sapoviruses and lagoviruses but may also represent a new calicivirus genus. The prevalence of these two BEC genotypes in cattle was unknown. Although reverse transcription-PCR (RT-PCR) primers for human NLV recognize NLV-BECs, the genetic relationships between NLV from humans and the NLV-BECs commonly circulating in cattle is undefined (Smiley et al., 2003).
To determine the genogroups and genotypes of bovine enteric caliciviruses (BECVs) circulating in calves, Myung et al. (2004) determined the complete capsid gene sequences of 21 BECVs. The nucleotide and predicted amino acid sequences were compared phylogenetically with those of known human and animal enteric caliciviruses. Based on these analyses, 15 BECVs belonged to Norovirus genogroup III and genotype 2 (GIII/2) and were genetically distinct from human Norovirus GI and GII. Six BECVs had capsid gene sequences similar to that of the unclassified Nebraska (NB)-like BECV. The 15 bovine noroviruses (BoNVs) were more closely related to Bo/NLV/Newbury-2/76/UK (GIII/2) and other known genotype 2 BoNVs than to genotype 1 Bo/NLV/Jena/80/DE. The BoNV Bo/CV521-OH/02/US showed high nucleotide and amino acid identities (84 and 94%, respectively) with the capsid gene of Bo/NLV/Newbury-2/76/UK, whereas the nucleotide and amino acid sequences of the RNA polymerase gene were more closely related to those of Bo/NLV/Jena/80/DE (77 and 87% identities, respectively) than to those of Bo/NLV/Newbury-2/76/UK (69 and 69% identities, respectively), suggesting that Bo/CV521-OH/02/US is a genotype 1-2 recombinant. Six NB-like BECVs shared 88 to 92% nucleotide and 94 to 99.5% amino acid identities with the NB BECV in the capsid gene. The results of this study demonstrated genetic diversity in the capsid genes of BECVs circulating in Ohio veal calves, provide new data for coinfections with distinct BECV genotypes or genogroups and describe the first natural BoNV genotype 1-2 recombinant, analogous to the previously reported human norovirus recombinants (Myung et al., 2004).
In summary, although calicivirus infections were confirmed in this area, but more detailed epidemiological studies and phylogenetic analysis of viruses isolated from more diverse geographical locations would be required in order to determine whether calicivirus is truly endemic in the Iran.
ACKNOWLEDGMENTS
We thank Dr. S.L. Oliver and Dr. A.M. Dastjerdi for valuable advisements and kindly supplying the primers and positive samples.
REFERENCES
- Oliver, S.L., A.M. Dastjerdi, S. Wong, L. El-Attar, C. Gallimore, D.W.G. Brown, J. Green and J.C. Bridger, 2003. Molecular characterization of bovine enteric caliciviruses: A distinct third genogroup of noroviruses (Norwalk-like viruses) unlikely to be of risk to humans. J. Virol., 77: 2789-2798.
Direct Link - Mead, P.S., L. Slutsker, V. Dietz, L.F. McCaig and J.S. Bresee et al., 1999. Food-related illness and death in the United States. Emerg. Infect. Dis., 5: 607-625.
Direct Link