
Hossam H. Abbas
Department of Hydrobiology, Veterinary Researches Division, National Research Center,
El-Bohooth St. Dokki, Giza, P.O. Box 12622, Cairo, Egypt
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 12 | Page No.: 2215-2221







ABSTRACT
The toxicity (LC50) of total and un-ionized ammonia was tested on different sizes (5, 10, 15 g) of common carp Cyprinus carpio maintained at three different pH levels within the range of 6.5-8.5. Within the same pH, the toxic effect of ammonia was independent of fish size. In addition, the toxicity of un-ionized ammonia increased at lower pH. However, total ammonia exerts some measure of toxicity and/or increased H+ concentration increases the toxicity of un-ionized ammonia. Common carp fingerlings (15 g) were exposed to the 96 h LC50 of un-ionized ammonia (0.93 mg NH3-N/L) at pH 7.5. Changes in hemoglobin (Hb), hematocrit (Ht), sodium (Na+), potassium (K+), glucose, aspartate aminotransferase (AST), alanine aminotransferase (ALT) and lactate dehydrogenase (LDH) were recorded. Blood Hb decreased after 6 and 96 h of exposure while Ht decreased after 3 h of exposure. Serum Na+ and K+ increased during the experimental period (96 h). Glucose concentration increased initially and then returned to less than the control value after 96 h of exposure. There was a significant decrease in AST, ALT and LDH activities after 6 h of exposure, after which enzyme activity increased until the end of the experimental period.
PDF Abstract XML References Citation
How to cite this article
Hossam H. Abbas, 2006. Acute Toxicity of Ammonia to Common Carp Fingerlings (Cyprinus carpio) at Different pH Levels. Pakistan Journal of Biological Sciences, 9: 2215-2221.
DOI: 10.3923/pjbs.2006.2215.2221
URL: https://scialert.net/abstract/?doi=pjbs.2006.2215.2221
DOI: 10.3923/pjbs.2006.2215.2221
URL: https://scialert.net/abstract/?doi=pjbs.2006.2215.2221
INTRODUCTION
Ammonia occurs in natural water in un-ionized (NH3) and ionized (NH4+) forms and can be a serious toxicant to fishes and other aquatic species. It enters water systems from several sources including industrial wastes, sewage effluent, agricultural input and animal feedlots. It is also a metabolic by-product of fish. The accumulation of ammonia in water used for intensive fish culture is a potential problem because it is toxic to fish. Most nitrogen in feeds and fertilizers that is not converted to fish flesh enters the water as ammonia, either by direct excretion from fish or by bacterial action on wastes. Ammonia concentrations can increase rapidly when water exchange rates are low (Alabaster and Lloyd, 1980; Harry and Boyd, 1987).
The toxicity of ammonia to different fish species has been extensively investigated (Thurston et al., 1978; Chatty et al., 1980; Russo, 1985; Bader, 1990; Abel, 1998; Salah El-Deen, 1999; Sampaio et al., 2002; Wicks and Randall, 2002; Chew et al., 2003). Ammonia toxicity depends principally upon the presence of NH3, which can readily diffuse across the gill membrane due to its lipid solubility and lack of charge, whereas the ionized form cannot readily pass through the hydrophobic micropores in the gill membrane (Sheehan and Lewis, 1986). However, NH4+ is excreted across the gill only via a carrier mediated process in exchange for Na+ and may also show considerable toxicity at low pH (Yamagata and Niwa, 1982; Chew et al., 2003).
The acute criterion for ammonia is dependent on pH and fish species and the chronic criterion is dependent on pH and temperature (USEPA.,1999). At lower temperatures, the dependency of chronic criterion is also dependent on the presence or absence of early life stages of fish (USEPA., 1999). Ammonia criteria identify pH as an important factor affecting the toxicity of ammonia and use an empirical model to describe the pH dependence of ammonia toxicity when expressed in terms of un-ionized ammonia (USEPA, 2002).
Sheehan and Lewis (1986) noted that lethal concentrations at pH = 6 were associated with very high total ammonia concentrations (2,000 mg N/L) and exhibited steeper concentration-effect curves than at higher pH. They also reported that other salts were lethal at similar concentrations and suggested that the toxicity of ammonia at low pH was due to the effect of osmotic shock on unacclimated organisms rather than a specific action of ammonium ion per se.
Because of the importance of un-ionized ammonia, it became a convention in the scientific literature to express ammonia toxicity in terms of un-ionized ammonia and water quality criteria and standards follow this convention. However, ammonium ion may contribute significantly to ammonia toxicity (Boyd, 1990). Observations that ammonia toxicity is relatively constant when expressed in terms of un-ionized ammonia come mainly from toxicity tests conducted at pH>7.5. At lower pH, toxicity varies considerably when expressed in terms of un-ionized ammonia and under some conditions is relatively constant in terms of ammonium ion (Erickson, 1985). Also, studies have established that mechanisms exist for the transport of ammonium ion across gill epithelia (Wood, 1993), so this ion might contribute significantly to ammonia exchange at gills and affect the accumulation of ammonia in blood, if the external concentration is sufficiently high.
The common carp Cyprinus carpio is an important fish for aquaculture in Egypt as one partial solution for meeting the increasing demand for protein. It has been artificially reproduced and cultured under Egyptian conditions. In view of the expanding culture of common carp, increasing numbers of hatcheries and lack of information on ammonia toxicity and its physiological effects on this species, the present study was undertaken to measure the acute toxicity of ammonia to different sizes of common carp at different pH levels. Another goal of the study was to define some physiological alterations in common carp fingerlings of stockable size and at the normal pH of Egyptian waterways (˜ 7.5), in response to acute exposure to the lethal concentrations of ammonia.
MATERIALS AND METHODS
Two experiments were conducted in May-July 2005 at the Central Laboratory for Aquaculture Research (CLAR), Abbassa fish farm, Abu-Hammad, Sharkia governorate, Egypt. The first determined the LC50 of un-ionized ammonia and ionized ammonia at different pH levels and fish sizes (3x3 factorial design). The second experiment assessed the hematological and physiological changes in common carp fingerlings (15 g) exposed to the 96 h LC50 of un-ionized ammonia.
Common carp were collected from Abbassa fish farm and individually examined for skin lesions or furunculosis and then acclimated to laboratory conditions for one week prior to the experiment. Each group of fish used in each test was the same size. Mortality was less than 5% during the acclimation period. Fish were stocked in test aquaria with dechlorinated aerated tap water. Fish were fed once daily with a 25% protein commercial pelleted fish diet (1 mm diameter) at 3% of body weight. Feeding was discontinued 48 h prior to and during the study. The experimental set-up consisted of thirty 112.5 L aquaria.
| Table 1: | Characteristics of the test water used in the experiment |
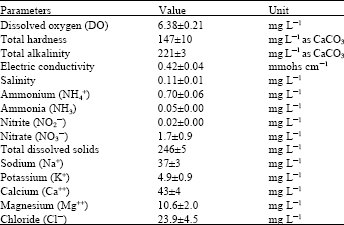 | |
| Data are presented as mean of three samples±SEM | |
Water was changed daily with water of the same specifications as each test aquarium to avoid fish metabolite accumulation. Aeration was discontinued during the experiment.
Reagent-grade ammonium chloride (Merck Company) and was mixed with tape dechlorinated water to obtain the required concentration. Dilute solutions of reagent-grade sodium hydroxide or hydrochloric acid were used to maintain the desired pH in test water. Before the start of each test, fish were placed in the test aquaria in water of pH 7.5. Adjustments in pH to the desired test pH conditions were made gradually and monitored manually over 24 h. Fish were maintained at the test pH for 48 h prior to starting the ammonia exposure test. Concentrations of un-ionized ammonia were calculated by means of the dissociation constants of Emerson et al. (1975) after measuring water temperature and pH. Ammonia concentration in each test aquaria was monitored daily by the phenol hypochlorite method (Solorzano, 1969) and concentrations were adjusted to initial levels by adding ammonium chloride or by partially exchanging water to remove ammonia when necessary.
Water samples characteristics were analyzed daily (Table 1) according to the method of APHA (1995).
In the toxicity test, fish were weighed individually and divided into three nominal weights (5, 10 and 15 g). Each group of fish was placed in three replicate aquaria for the same weight and for every studied concentration (12 fish/aquarium) and exposed to different concentrations of total ammonia at different pH levels. Fish in tests 1 to 3, 4 to 6 and 7 to 9 were challenged with ammonia at pH values of approximately 6.5, 7.5 and 8.5, respectively. The concentration-response relationship (LC50) and 95% confidence intervals (CI) were determined according to USEPA., (2002).
To assess the physiological effect of ammonia on common carp, fingerlings (15 g) were placed in six aquaria (12 fish/aquarium) with water containing the 96 h LC5o (0.93 mg NH3-N/L) determined from the first experiment. The pH of the solution was adjusted to 7.5 every 6 h by using sodium hydroxide or hydrochloric acid. A freshly prepared solution was replaced every 24 h of exposure. A control group with no toxicant was also included in the experiment and treated similarly.
Group of six fish were selected from test aquaria after 3, 6, 12, 24, 48 and 96 h of exposure to the toxicant. Blood samples were withdrawn from the caudal artery with a sterile syringe with a heparinized glass pipette inserted through the middle line just behind the anal fin in a dorso-cranial direction.
Hemoglobin content (Hb) was estimated by using the cyanmethemoglobin method described by Boehringer Mannheim kit according to Zijlstra (1961). Hematocrit (Ht) was determined in small heparnized hematocrit capillary tubes, with a hematocrit centrifuge at 3000 rpm for 15 min. Serum glucose concentration was measured by using the GOD-PAP method (enzymatic colorimetric method) according to Trinder (1969). Serum electrolytes (Na+ and K+) were measured by atomic absorption (Perkin Elmer 2280) according to Fernandez and Khan (1971). Serum AST and ALT activities were determined colorimetrically by transaminase kits according to Reitman and Frankel (1957). Serum lactate dehydrogenase (LDH) activity was determined by the enzymatic reaction.
Data in the first experiment were analyzed with a two-way analysis of variance. In the second experiment, data were analyzed with a one-way analysis of variance to identify the significant differences among sampling points in the control and treated groups. The F-test was used to compare between treatments and the control group (p≤0.05). All statistical tests performed using statistical analysis software (SAS, 2000).
RESULTS AND DISCUSSION
Ammonia toxicity: There were no significant differences in ammonia toxicity among different fish sizes at each of the three pH values (Table 2). This indicates that the toxic effect of ammonia is independent of carp size across the range 5-15 g. Dosdat et al. (2003) found that the final average weights of sea bass subjected to different concentrations of un-ionized ammonia and total ammonia nitrogen were similar in all treatments. However, Thurston and Russo (1983) reported that sensitivity varied in rainbow trout with greater sensitivity in late alevin and senescent adults.
When pH increases, ammonia toxicity increases. If the un-ionized ammonia is solely responsible for the toxic action on the test fish, then one would expect that the LC5o values for un-ionized ammonia would be reasonably constant for all tests regardless of the solution pH and total ammonia present, but this was not the case. The maximum LC5o values in terms of un-ionized ammonia occurred at pH 8.5, while the values at pH 6.5 are only 43% of these or less (after 96 h of exposure). Un-ionized ammonia LC50 values are markedly less at low pH, than they are at high pH values. Thus the toxicity of NH3 is not constant over the pH range tested, i.e. increased H+ concentration increases the toxicity of NH3. This explanation is consistent with the work of Thurston et al. (1981, 1983, 1984, 1986), Salah El-Deen (1999), USEPA. (1999, 2002) and Chew et al. (2003). Thus, the toxicity of ammonia to common carp in terms of un-ionized ammonia alone does not remain constant over the pH range considered acceptable to freshwater fishes.
Effect of ammonia exposure on serum constituents: Hemoglobin content (Hb) of common carp exposed to the 96 h LC50 of ammonia showed a significant change when compared to the control value (Fig. 1A). However, only after 6 and 96 h exposure was a significant decrease in Hb content recorded compared to the control value. In contrast, the Ht level significantly decreased directly after 3 h of exposure and continued till the end of the experiment period (96 h).
Studies of blood parameters in fishes subjected to conditions of elevated environmental ammonia have resulted in conflicting conclusions. No changes were observed in the number of erythrocytes, Hb or Ht in the freshwater Labeo capensis upon short exposure to sublethal (Hattingh, 1976) and lethal (Smart, 1978) levels of ammonia. Whereas Ahmed et al. (1992) reported an initial increase in erythrocytes, Hb and Ht levels of Oreochromis niloticus subjected to a 24 h exposure to 16.6% of 96 h LC50 of ammonia; the increase was followed by significant decrease in the same parameters until the end of the exposure time (7 days). The authors attributed the initial increase to the coexisting process of renovation of erythrocytes due to the release of immature erythrocytes from the spleen, while the decrease may reflect a hemolytic anemia. Another explanation was provided by Niels et al. (1998), who attributed the increase of Hb and Ht in rainbow trout Oncorhynchus mykiss after one day of ammonia exposure to the hemoconcentration via the release of large numbers of immature erythrocyte from the spleen to challenge the stress or increased diuresis.
In the present study, the decrease in Hb and Ht after acute exposure to ammonia could be attributed to shrinkage of erythrocytes, decrease of erythrocytes production in the hematopoietic tissue and hemodilution and intravascular destruction due to the acute toxicity of ammonia.
| Table 2: | Acute toxicity of un-ionized ammonia and ammonium (as N) to common carp at different pH and fish size |
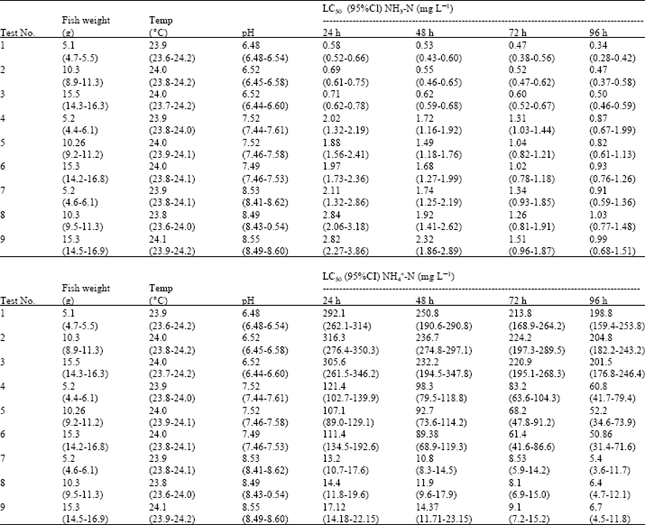 | |
Data reported for fish weight, temperature and pH are mean values of all measurements in all test aquaria with ranges in parentheses. Toxicity tests were determined from three replicates per trial with 36 fish per test. CI: Confidence Intervals | |
Similar results have been reported for the African catfish Clarias gariepinus and blue tilapia Oreochromis aureus exposed to lead and copper (El-Nagar et al., 2001), mercury (Mazhar et al., 1987a); crude oil (Mazher et al., 1987b) and for Onchorhyncus kisutch (Buckley et al., 1979); Oreochromis niloticus (Ahmed et al., 1992) exposed to toxic concentration of ammonia and Ctenopharyngodon idella exposed to acute toxicity of ammonia (Salah El-Deen, 1999).
Serum glucose levels significantly increased after 3 h of exposure to ammonia. However, serum glucose level decreased after 24 h of exposure until the end of exposure time (96 h). The results of the studied enzyme activities in the serum of common carp showed that there was only a significant decrease in AST, ALT and LDH activities at 6 h of exposure, after which the enzyme activities significantly increased till the end of the experimental period (Fig. 1B).
Ammonia intoxication caused initial significant increase in glucose concentrations indicating that the common carp experienced physiological stress during the first 12 h of exposure, followed by a decrease in glucose concentration to near the control value, indicating that the fish reestablished homeostasis (Selye, 1973). A similar effect was measured in rainbow trout Onchorhyncus mykiss exposed to 0.34 mg NH3/L (Thurston et al., 1978) and in grass carp Ctenopharyngodon idella exposed to 0.7 mg NH3/L at pH 7.5 (Salah El-Deen, 1999).
Serum Na+ increased during the entire experimental period except after 24 h of exposure. Whereas serum K+ showed an increase at all the exposure times and was pronounced (p<0.05) after 3, 24 and 48 h of exposure (Fig. 1C).
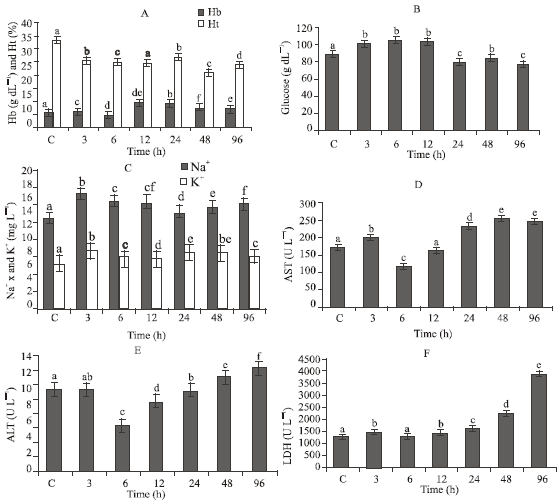 | |
| Fig. 1: | Levels of some physiological characteristics in common carp fingerlings (15 g body weight) subjected to the 96 h LC50 at pH 7.5. Bars with the same letter are not significantly different (p>0.05) and represented as Mean±SEM |
At elevated ambient ammonia concentration, movement of NH3 will be along the NH3 gradient between blood and water, whereas excretion of NH4+ will occur by exchange of NH4+ for external Na+ across the branchial cells (Maetz, 1973; Smith, 1982) on the basis of excretion of NH4+ or H+ in exchange for Na+ and HCO3- for Cl- across the gills (Maetz and Garcia Romeu, 1964; Smith, 1982). The significant elevation of Na+ in the studied fish (Fig. 1C) may therefore be explained by increased intake resulting from accelerated excretion of NH4+ to reduce serum ammonia. However, the slight reduction in Na+ after 24 h of acute exposure to ammonia is consistent with the mechanism of competition between external NH4+ and Na+ for common entry sites through the gills when excretion occurs against a concentration gradient (Maetz, 1973; Salah El-Deen, 1999; Chew et al., 2003).
The fluctuation of K+ concentration in the serum of common carp in the present experiment may have resulted from intravascular damage to erythrocyte membrane and subsequent leaking of K+ into serum, reduced inward transport, or some combination of these factors. This assumption is in agreement with the work on coho salmon, Onchorhyncus kisutch by Buckley et al. (1979) and the proposed mechanism of hemolyses by ammonia (Heath, 1987).
The increased levels of sodium and potassium might be due to the renal dysfunction (Lauran and McDonald 1985) and/or may be due to the alteration in the active transport of ions (Tulasi et al., 1990). The ionic disturbances could also be attributed to the outward leakage of intracellular ions, especially potassium, caused by stress (Haux and Larsson, 1982).
Stress acts by inhibiting certain enzymes, thus interfering with metabolic processes (Weis et al., 1981). In the present study, AST, ALT and LDH activities increased in serum of common carp after 3, 24, 48 and 96 h of exposure to ammonia (Fig. 1D-E). The increase in serum aminotransferase is indicative of some degree of tissue necrosis (Niels et al., 1998) or of liver and kidney dysfunction and leakage of these enzymes from injured tissue into the blood (Salah El-Deen and Rogers, 1993; Salah El-Deen, 1999). In addition, the increase in ALT activity might also be caused by the increased availability of pyruvate formed from increased LDH activity (Chatty et al., 1980). The increase in ALT activity corresponds to LDH activity. The increase level of AST and ALT in common carp after exposure to ammonia may also be due to the loss of Kreb’s cycle, with the result that these enzymes compensate by providing α-ketoglutarate (Chatty et al., 1980; Salah El-Deen, 1999). The observed changes could be also due to generalized organ system failure as the animals approach death due to the effect of ammonia. The physiological and biochemical changes expressed by fish in response to ammonia toxicity and water toxification are serious problems facing the culturists and researchers who are working in the field of aquaculture especially the intensive aquaculture system, it affect the fish health and subsequently the fish growth.
Thus, we concluded that ammonia toxicity is independent of common carp size and is not constant over the pH range (6.5-8.5) indicating that increased H+ concentration increases the toxicity of ammonia. The result of analyses of blood samples collected in the field can be used as an indicator of potential adverse affects on the survival, growth performance and reproductive success of fish.
REFERENCES
- Dosdat, A., J.P. Le Ruyet, D. Coves, G. Dutto, E. Gasset, A. Le Roux and G. Lemarie, 2003. Effect of chronic exposure to ammonia on growth food utilization and metabolism of the European sea bass Dicentrarchus labrax. Aquatic Living Resour., 16: 509-520.
CrossRef - Harry, V.D. and C.E. Boyd, 1987. Acute toxicity of ammonia and nitrite to spotted sea trout. Prog. Fish Culturist, 49: 260-263.
Direct Link - Hattingh, H.J., 1976. Blood sugar as an indicator of stress in the freshwater fish, Labeo capensis (Smith). J. Fish. Biol., 10: 191-195.
CrossRefDirect Link - Haux, C. and A. Larsson, 1982. Influence of inorganic lead on the biochemical blood composition in the rainbow trout, Salmo gairdneri. Ecotoxicol. Environ. Saf., 6: 28-34.
CrossRefPubMedDirect Link - Lauran, D.J. and D.G. McDonald, 1985. Effects of copper on branchial ionregulation in the rainbow trout, Salmo gairdneri Richardson. J. Comp. Physiol., 155: 635-644.
CrossRef - Niels, E.V., B. Korsgaard and F.B. Jenson, 1998. Isolated and combined exposure to ammonia and nitrite in rainbow trout Oncorhynchus mykiss effects on electrolyte status, blood respiratory properties and brain glutamine glutamate concentrations. Aquatic Toxicol., 41: 325-342.
Direct Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Salah El-Deen, M.A. and W.A. Rogers, 1993. Changes in total protein and transaminase activities of grass carp exposed to diquat. J. Aqua. Anim. Health, 5: 280-286.
CrossRef - Sampaio, L.A., W. Wasielesky and K.C. Miranda-Filho, 2002. Effect of salinity on acute toxicity of ammonia and nitrite to juvenile Mugil platanus. Bull. Environ. Contamin. Toxicol., 68: 668-674.
CrossRef - Sheehan, R.J. and W.M. Lewis, 1986. Influence of pH and ammonia salts on ammonia toxicity and water balance in young channel catfish. Trans. Am. Fish. Soc., 115: 891-899.
CrossRefDirect Link - Solorzano, L., 1969. Determination of ammonia in natural waters by the phenolhypochlorite method 1 1 This research was fully supported by U.S. atomic energy commission contract No. ATS (11‐1) GEN 10, P.A. 20. Limnol. Oceanogr., 14: 799-801.
CrossRefDirect Link - Thurston, R.V., R.C. Russo and C.E. Smith, 1978. Acute toxicity of ammonia and nitrite to cutthroat-trout fry. Trans. Am. Fish. Soc., 107: 361-368.
Direct Link - Thurston, R.V., R.C. Russo and G.A. Vinogradov, 1981. Ammonia toxicity to fishes effect of pH on the toxicity of un-ionized ammonia species. Environ. Sci. Technol., 15: 837-840.
CrossRef - Thurston, R.V., R.J. Russo, R.J. Luedtke, C.E. Smith and E.L. Meyn et al., 1984. Chronic toxicity of ammonia to rainbow trout. Trans. Am. Fish. Soc., 113: 56-73.
Direct Link - Wicks, B.J. and D.J. Randall, 2002. The effect of feeding and fasting on ammonia toxicity in juvenile rainbow trout Oncorhynchus mykiss. Aqua. Toxicol., 59: 71-82.
Direct Link