
Farouk Al-Quadan
College of Sciences, Department of Biological Sciences, Hashemite University, P.O. Box 150459, Postel Code 13115 Zarqa, Jordan
Fawzia M.R. Al-Charchafchi
College of Sciences, Department of Biological Sciences, Hashemite University, P.O. Box 150459, Postel Code 13115 Zarqa, Jordan
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 2 | Page No.: 210-213







ABSTRACT
Germination of Artemisia herba alba seeds in the presence of esculetin showed lower G6PDH or 6PGDH activities compared to that germinated in esculetin absence; however, the same enzymes showed higher activities upon germination in the dark compared to that under light. Both enzymes were continuously increased from day one to day five during germination. Of the isoenzyme pattern found G6PDH or 6PGDH, the fast moving band of G6PDH or the slower moving band of 6PGDH was influenced by germination under dark condition or presence of esculetin.
PDF Abstract XML References Citation
How to cite this article
Farouk Al-Quadan and Fawzia M.R. Al-Charchafchi, 2006. G6PDH and 6PGDH Activities of Artemisia herba alba Seeds and Seedlings During Germination in Presence of Esculetin. Pakistan Journal of Biological Sciences, 9: 210-213.
DOI: 10.3923/pjbs.2006.210.213
URL: https://scialert.net/abstract/?doi=pjbs.2006.210.213
DOI: 10.3923/pjbs.2006.210.213
URL: https://scialert.net/abstract/?doi=pjbs.2006.210.213
INTRODUCTION
Artemisia herba alba is a common perennial shrub in the semi-deserts of the Middle East (Zohary, 1973); it is one of the most important medicinal species of Artemisia (Al-Charchafchi and Jawad, 1982a, b).
Phenolic compounds were found to be phytotoxic constituents of many plant species extracts (An et al., 2000); moreover, were elevated in plants under stress conditions (Shetty et al., 2002; Randhir and Shetty, 2005). Phenolic compounds caused seeds dormancy, inhibit seed germination and suppress photosynthesis in Artemisia herba alba (Al-Charchafchi et al., 1987; Alam et al., 2001; Muscolo et al., 2001). Erythrose-4-phosphate, an intermediate of pentose phosphate pathway, can combine with phosphoenol pyruvate to initiate shikimic acid pathway; which is used by many plants in the biosynthesis of their phenolic compounds (Ashihara and Komamine, 1964). Glucose-6-phosphate dehydrogenase (G6PDHE.C1.1.1.49) and 6 phosphogluconate dehydrogenase (6PGDH E.C 1.1.1.44) are the key enzymes of pentose phosphate pathway (Andrei et al., 2002; Sükrü et al., 2003).
The hypothesis under investigation in this study was the role played by the phenolic compound esculetin in regulating pentose phosphate pathway activity in Artemisia herba alba during seed germination. This is accomplished by studding G6PDH and 6PGDH activities in the presence and absence of esculetin.
MATERIALS AND METHODS
Plant material: Seeds of Artemisia herba alba used in the study were harvested at December 2003 from the medicinal and aromatic plant garden of the Hashemite University/Zarka, Jordan. The seeds were kept in brown paper bags in refrigerator until used.
Germination and growth: Germination was performed in 12 cm diameter petri dishes lined with one layer of filter paper (Whateman No. 30) and moistened with 5 mL of distilled water or esculetin at 0.2 and 0.4 mM. 100 seeds per dish (three replicates) were used for each treatment. All dishes were incubated at 20°C in growth chamber under continuous illumination or darkness for 1, 3 and 5 days. Dark treatment was conducted by wrapping the dishes with aluminum foil immediately after seed imbibition. Esculetin was prepared just prior to use and was wrapped in aluminum foil container.
G6PDH and 6PGDH extraction procedure: Extraction solution consisted of 100 mM Tris-HCl (pH 7.6), 2 mM EDTA Na2 [Scharlau, Spain] and 30 mM β-Mercaptoethanol [ARCOS, USA].The imbibed seeds and seedlings were suspended in 1 mL of extraction solution and were homogenized at 25000 rpm for 3 min using IKA homogenizer [Germany]. During homogenization samples were immersed in ice bath. Homogenates were then centrifuged for 20 min at 10000 rpm at 4°C; supernatant was re-centrifuged two more times for 5 min to remove lipid debris.
| Table 1: | Artemisia herba alba seeds G6PDH specific activities U/L/mg after one, three and five days of growth in presence and absence of different concentrations of esculetin under light and dark conditions |
 | |
| Table 2: | Artemisia herba alba seeds 6PGDH specific activities U/L/mg after one, three and five days of growth in presence and absence of different concentrations of esculetin under light and dark conditions |
 | |
Supernatants were used immediately for G6PDH and 6PGDH activity determination, or stored at -70°C until time for analysis.
G6PDH and 6PGDH assays: The assay procedure of Hohorst (1965) was used with slight modification. All assays were performed with GENSYS 5, USA spectrophotometer. Each assay consisted of 850 μL Tris-HCl buffer (10 mM, pH 7.6), 100 μL MgCl2 (67 mM), 10 μL glucose-6-phosphate or 6-Phosphogluconate (10 mM) [Sigma, Mo, USA] and 10 μL NADP+ (5 mM) [BDH, England] in a total volume of 1 mL.
Protein determination: Quantitative protein determination by the method of Bradford (1976), with Bovine Serum Albumin (BSA) was used as standard.
Electrophoresis: Polyacrylamide gel electrophoresis was carried out as described by Ornstein and Davis (1962) using 7.5% acrylamide and Bio-Red vertical cell (USA). Isoenzymes were visualized according to procedure described by Schnarrenberger et al. (1973). The gels were photographed using digital camera (JAI, Japan) interfaced to a computer (DELL, USA).
Statistical analysis: ANOVA test was used to determine the level of significance within the Artemisia herba alba species regarding the effect of esculetin on G6PDH and 6PGDH activities. Significance of differences was accepted when p< 0.05.
RESULTS
G6PDH and 6PGDH activities were increased continuously from day 1 to day 5 upon seed imbibition and growth under light or darkness (p<0.05). On the other hand G6PDH and 6PGDH activities were significantly decreased (p<0.05 ) , in the presence of 0.2 or 0.4 mM esculetin, in comparison with activities in the absence of esculetin under light or darkness (Table 1 and 2).
G6PDH isoenzymes: After one or three days of seed germination and growth under light or in the presence of 0.2 mM esculetin, electrophoresis study showed one G6PDH isoenzyme band (Rf 0.52); after three days of growth and in the presence of 0.4 mM esculetin however, an additional slower isoenzyme band (Rf 0.49) was observed with lower intensity compared to the more intense fast moving band mentioned.
After five days of growth in the presence or absence of 0.2 and 0.4 mM esculetin and under light or dark condition, three isozymes were observed as shown in Fig. 1; 1-7; dark germination (in the presence or absence of 0.2 or 0.4 mM esculetin) showed igher intensity for the fast moving band Rf 0.52 compared with slower band (Rf 0.49) as shown in Fig. 1 lane 8.
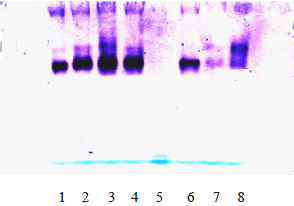 | |
| Fig. 1: | G6DH isoenzymes pattern during seed germination of Artemisia herba alba . Lane 1,6 in 0.4 mM esculetin for 5 days under dark; Lane 2, in 0.2 mM esculetin for 5 days under dark; Lane 3, in distilled water for 5 days under dark; Lane 4, in distilled water for 5 days under dark; Lane 7, in 0.2 mM esculetin for 5 days under light; Lane 8, in distilled water for 5 days under light |
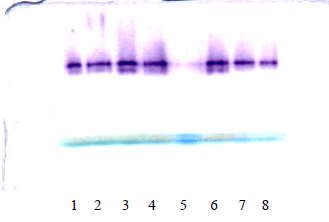 | |
| Fig. 2: | Expression of 6PGDH isoenzymes during seed germination of Artemisia herba alba. Lane 1, in 0.4 mM esculetin for 5 days under dark; Lane 2, in 0.2 mM esculetin for 5 days under dark; Lane 3, in distilled water for 5 days under dark; Lane 4, in distilled water for 5 days under dark; Lane 6, in 0.4 mM esculetin for 5 days under light; Lane 7, in 0.2 mM esculetin for 5 days under light; Lane 8, in distilled water for 5 days under light |
6PGDH isoenzymes: After 1, 3 and 5 days of seed germination under light condition, one 6PGDH isoenzyme at Rf (0.50) was observed; however, seed germinated in the presence of 0.4 mM esculetin and under light condition showed additional fast moving band (at Rf 0.53) as shown in Fig. 2 lane 6; this additional band was observed in seeds grown in dark in the presence of distilled water Fig. 2 lane 3. The slower band had higher relative intensity compared to the fast moving band.
DISCUSSION
Artemisia herba alba G6PDH and 6PGDH activities were gradually increased from day one to day five of germination. These results were in agreement with previous results (Randhir et al., 2004; Mitchell and Barrett 2001; Mitchell et al., 1999).
This increase in activity may be due to switch from glycolysis to pentose phosphate pathway (Mayer and poljakoff-Mayber, 1989) or to fulfill the increased demand for NADPH associated with growth (Bewley and Black, 1994).
In seed germinated in the dark, the elevated activities of G6PDH and 6PGDH found in this study, were to compensate the short supply of NADPH2 resulted from the halt of photosynthetic electron flow (Robinson, 2000). Randhir and Shetty, (2005) indicated the differential expression pattern between cytosolic and plastidic isoforms in spinach and soybean plants germinated in dark; as oxidative portion of the pentose phosphate pathway is found in both cytosolic and plastidic fractions of photosynthetic cells (Perry and Shetty, 1999).
Presence of esculetin during germination of seeds and seedlings of Artemisia herba alba, have influenced both activities and isoenzymes patterns of G6PDH and 6PGDH as judged by band intensities; these results elucidated the role played by esculetin phenolic compound in regulating pentose phosphate pathway activity in Artemisia herba alba during seed germination. Other explanations based on root and shoot isoenzymes cannot be ruled out (Mitchell and Barrett, 2001).
These results were consistent with earlier study on pentose phosphate pathway enzymes from Nicotiana tabacum (Hoover et al., 1977) were esculetin was found to be inhibitory effect on one band and stimulatory effect on the other band.
This study has elucidated the role played by esculetin phenolic compound in regulating pentose phosphate pathway activity in Artemisia herba alba during seed germination.
REFERENCES
- Alam, S.M., S.A. Ala, R. Ansari and M.A. Khan, 2001. Influence of leaf extract of bermuda grass (Cynodon dactylon L.) on the germination and seedling growth of wheat. Wheat Inform. Service, 92: 17-19.
Direct Link - Andrei, L.K., H. Ji-Biao, C. Tinnakorn, M.F. Ryan, M.K. Yeon, R. Roberto and R.P. Howard, 2002. Pregnancy alters glucose-6-phosphate dehydrogenase trafficking, cell metabolism and oxidant release of maternal neutrophils. J. Clin. Invest., 110: 1801-1811.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Randhir, R., Y. Lin and K. Shetty, 2004. Phenolics, their antioxidant and antimicrobial activity in dark germinated fenugreek sprouts in response to peptide and phytochemical elicitors. Asia Pac. J. Clin. Nutr., 13: 295-307.
Direct Link - Randhir, R. and K. Shetty, 2005. Developmental stimulation of total phenolics and related antioxidant activity in light- and dark-germinated corn by natural elicitors. Process Biochem., 40: 1721-1732.
Direct Link - Shetty, P., T.M. Atallah and K. Shetty, 2002. Effects of UV treatment on the proline-linked pentose phosphate for phenolics and L-DOPA synthesis in dark germinated Vicia faba. Process Biochem., 37: 1285-1295.
Direct Link