
Erdal Balcan
Molecular Biology Section, Department of Biology, Faculty of Science and Art, Celal Bayar University, 45047 Muradiye Campus, Manisa-Turkey
Mesut Sahin
Molecular Biology Section, Department of Biology, Faculty of Science and Art, Celal Bayar University, 45047 Muradiye Campus, Manisa-Turkey
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 10 | Page No.: 1852-1860







ABSTRACT
During the maturation stages of thymocytes, apoptosis plays a crucial role for the production of immunocompetent T cells. On the other hand, changes of glycosylation may influence both T cell signaling and T cell development. Carbohydrate molecules are not only energy sources of the cells, but they can also responsible for the recognition phenomena such as protein targeting and cell to cell interactions. Therefore, glycosylation of cell surface proteins or lipids controls critical T cell processes, such as T cell death. Sialic acids, terminal sugar of the glycan motifs, are primarily responsible for the cellular interactions. In the present study, we purposed to determine of the cell surface carbohydrate alterations and the possible relationships of apoptosis with these alterations in the thymocytes from different periods of embryonic and newborn thymic tissues. Our terminal deoxynucleotidyltransferase-mediated dUTP nick end labeling (TUNEL) staining indicated that the thymocyte apoptosis is increased after the day 16of embryonic development and it proceeds in neonatal period. In addition, the surface density of sialic acids were decreased in newborn thymi when compared to embryonic tissues. These results indicated that, there is a significant relationship between cell surface sialic intensities and apoptosis. Thus, in the embryonic development, pathways which responsible for the thymocyte apoptosis may different from those of the newborn period due to masking effects of sialic acids. However, after the birth, sialic acid intensities were highly diminished in the thymocytes and, therefore they may undergo to apoptosis by another pathway(s), such as galactin-1 induced pathway.
PDF Abstract XML References Citation
How to cite this article
Erdal Balcan and Mesut Sahin, 2006. Apoptosis-Sialic Acid Relationships at Different Developmental Stages of Murine Thymus. Pakistan Journal of Biological Sciences, 9: 1852-1860.
DOI: 10.3923/pjbs.2006.1852.1860
URL: https://scialert.net/abstract/?doi=pjbs.2006.1852.1860
DOI: 10.3923/pjbs.2006.1852.1860
URL: https://scialert.net/abstract/?doi=pjbs.2006.1852.1860
INTRODUCTION
The thymus gland is an important organ for the maturation and selection of antigen-specific, self-tolerant T lymphocytes from marrow derived precursors and the first morphological events of thymus organogenesis are seen between days 10 and 13.5 of embryonic development (Manley and Blackburn, 2003). During the maturation stages, thymocytes exhibit characteristic phenotypes. The earliest thymocytes lack functional TCR as well as CD4 and CD8 coreceptor expression, but can be identified by combinations of cell surface receptors, CD44 and CD25. They classified as DN1 (CD44+ CD25-), DN2 (CD44+ CD25+), DN3 (CD44- CD25+) and DN4 (CD44- CD25-) (Zacharchuk et al., 1991; Page et al., 1996; Godfrey et al., 1993). In the maturation of immunocompetent T cells, the interaction of developing thymocytes with thymic epithelial cell is an important step (Singer and Haynes, 1987; Patel and Haynes, 1993). CD4+ and CD8+ thymocytes expressing αβ T-cell receptor (TCR) must interact productively with self-peptide-major histocompatibility complex (MHC), a process termed positive selection (Anderson et al., 1999). However, they will deleted by apoptosis as a consequence of a TCR that cannot interact with self-peptide-MHC complexes (death by neglect) or one that binds to these complexes with high affinity, a process termed negative selection (Sebzda et al., 1999; Minter and Osborne, 2003, Smith et al., 1989; Nossal, 1994; Sprent and Kishimoto, 2002). Apoptosis is a crucial event during the T lymphocyte development for the production of immunocompetent T cells (Cohen et al., 1992).
Apoptosis, or programmed cell death, is a mechanism for killing of no longer needed, damaged, infected or dangerous cells (Kerr et al., 1972). This mechanism is a crucial event during the T lymphocyte development for the production of immunocompetent T cells (Cohen et al., 1992). There are two main pathways in apoptotic mechanisms: The intrinsic cell death pathway controlled by Bcl-2 family members and the extrinsic cell death pathway initiated by ligation of a trans-membrane death receptor of the tumor necrosis factor receptor type 1 superfamily. A particular member of this family, Fas (also known as CD95 or APO-1), is an integral membrane protein that plays a unique role in apoptosis of lymphoid cells (Strasser, 1995; Nagata, 1997; Wallach et al., 1998). These two pathways work together to regulate T lymphocyte development and function (Zhang et al., 2005).
Cell surface carbohydrate molecules, especially sialic acids, serve as molecular determinants for mediating in many cellular interactions (Ofek and Sharon, 1990; Varki, 1992, 1997; Feizi, 1993). Sialic acids are nine-carbon carboxylated monosaccharides. Due to their location at the terminal position on glycans of cell surface glycoproteins and glycolipids and to net negative charge at physiological pH, they serve as a potentially regulator of the cellular interactions (Varki, 1997). Sialic acids play a dual role in recognition phenomenon: in addition to antigenic effects, they are also responsible for the masking of antigenic and other recognition sites of soluble molecules and cells (Ashwell and Harford, 1982; Schauer 1982, 1985; Schauer et al., 1984).
A number of reports have demonstrated that the cell surface sialylation changes as thymocytes mature (Gillespie et al., 1993; Wu et al., 1997; Krishna and Varki, 1997; Despont et al., 1975; Reisner et al., 1976; Akashi et al., 1988; Whiteheart et al., 1990; Baum et al., 1996) and regulation of glycoprotein sialylation may control susceptibility to cell death at specific points during T cell development (Amano et al., 2005). During the maturation, the cell surface glycoconjugates of thymocytes display different sialylation patterns whose physiologic significance still remains unknown. Some endogenous lectins which participating to T cell maturation, such as galactin-1 (Baum et al., 1995; Hughes, 1992), CD22 (Sgroi et al., 1995), CD23 (Kijimoto-Ochiai and Ueda, 1995) and L-selectin (Varki, 1994) can bind unique carbohydrate ligands that require (CD22 and L-selectin) or are inhibited by (galactin-1 and CD23) the presence of sialic acids. Galactin-1 is a member of the family of β-galactoside binding proteins and responsible for the apoptosis of immature cortical thymocytes (Perillo et al., 1997). On the other hand, the alteration of cell surface sialylation leads to apoptosis of the CD4+CD8+ double positive thymocytes (Mucci et al., 2002).
The aim of this study was to evaluate the alterations in the sialylation pattern of thymocytes from embryos and newborn mice belonging the different age groups and to apoptosis and sialic acid relationships during the thymus organogenesis.
MATERIALS AND METHODS
Animals: 18 Balb/c mice (14 female, 4 male) were provided from Ege University Medical Faculty Animal Research Center, Izmir. In order to obtain inbred lines, mice were mated randomly from March to October 2005.
Preparation of tissues: Eight or ten week old inbred male and female Balb/c mice were caged overnight. The day the vaginal plug was found was designated embryonic day 0. Mice were scarified during their gestational periods at 14, 15, 16 and 17th days and embryos were extracted. For each time, at least five litters were examined. On the other hand, 1, 5 and 13 days old newborn mice were thymectomized. All of the embryos and thymi from newborns were fixed with Bouin’s solution and formalin, dehydrated and embedded to paraffin according to routine procedure and 5 μm sections were prepared.
Apoptosis assay: In order to determine of apoptotic cells, terminal deoxyribonucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assay were performed using Promega’s DeadEndTM Calorimetric TUNEL system (Cat # G7130) according to the manufacturer’s instructions. Briefly, following postfixation in 4% paraformaldehyde, permeabilisation with proteinase K treatment, 4% paraformaldehyde again and PBS washes, samples were incubated with biotynlated nucleotide and TdT enzyme at 37°C for 30 min. Following washes and blocking endogen peroxidase activity using H2O2, the biotyn labels were detected with streptavidin-HRP (horse radish peroxidase) and a DAB (diaminobenzidine tetrahydrochloride dihydrate) color reaction. For the positive control, after the second fixation with 4% paraformaldehyde, some tissues were treated with 1 U mL-1 RNAse free DNAse in separated Coplin jars.
Histochemistry of tissues: All thymic tissues used in this study were stained histochemically with a unique physical development procedure for the detection of sialic acid moieties. This method known as Selective periodate oxidation-Phenylhydrazina-Thiocarbohydrazide-Silver protein-Physical development and based on selective periodate oxidation of sialic acid residues yields C7 monoaldehydes, which condense with phenylhydrazine to give rise to corresponding phenylhydrazone. This can undergo solvolysis to yield TCH-SP-PD-reactive monoaldehydes (Ueda et al., 1995). After the a saponification technique (Culling et al., 1974), for the removal of O-acyl esters from sialic acids, tissues were oxidized with periodic acid and immersed in 1% phenylhydrazonium chloride aqueous solution. Following the washing period, tissues were incubated with 0.5% thiocarbohydrazide in 10% acetic acid for 40 min at 40°C and were immersed in 1% silver protein aqueous solution for 30 min in a dark box. Tissues were physically developed with a mixture of two solutions which contain 20% gum arabic with 10% silver nitrate and hydroquinone with citric acid, respectively. The working solution was prepared immediately before use. Finally t issues were rinsed a photographic fixer diluted 1:4 for 5 min. and dehydrated. The reaction products obtained by this method were colored brown and black. For the confirmation of staining method can specifically detect sialic acids, in combination with this method, we performed a neuraminidase digestion. Briefly some sections were treated with 1 U mL-1 Clostridium perfiringens neuraminidase (Type X, Sigma, Germany) which hydrolyzes α (2→3), α (2→6), α (2→8) glycosidic linkages of terminal sialic acid residues of various glycoconjugates, in 0.1M Na-acetate buffer containing 10 mmol L-1 CaCl2 for 18 h at 37°C before the staining procedure.
RESULTS
Apoptosis: With the TUNEL stainings it was seen that a few apoptotic cells are found in cortical regions of thymus from 14 and 15th embryonic days, however, after the day 16 of embryonic development high number of apoptotic cells were detected in not only cortex but also in corticomedullary region and medulla. When the compared with embryonic tissues, neonatal thymi contain many apoptotic cells in different regions (i.e., cortex, corticomedullar region and medulla). Especially after 5th day of birth, apoptotic thymocytes were increased in these regions (Fig. 1).
PA-P-TCH-SP-PD: When the tissue sections from thymic tissues in days 15, 16 and 17 of embryonic development and in days 1, 5, 9, 13 and 16 of newborn were stained with a specific technique, periodic acid-thiocarbohydrazide-silver protein-physical development (PA-P-TCH-SP-PD) procedure, which demonstrate the localization of sialic acids, the reaction intensities of embryonic cortical and medullar thymocytes were strong, but some thymocytes in the cortex and medulla were not stained. Very strong staining intensities were also detectable in the capsules and intracellular spaces of thymic stroma. The digestion with Clostridium perfiringens Type X strongly diminished the intensities of PA-P-TCH-SP-PD reactions of thymocytes from 17th embryonic day (Fig. 2).
On the other hand, the PA-P-TCH-SP-PD reaction was evidently decreased in 1, 5 , 9, 13 and 16th neonatal days. Only a few thymocytes were stained in the cortex and medulla. This indicated that at least two different physiological situation may occur in neonatal thymus (Fig. 3).
DISCUSSION
The thymus gland, is a primer lymphoid organ, have a central role in the maturation and selection of thymocytes. The thymus consist of a cortex, containing immature thymocytes and a medulla, containing mature thymocytes that are derived from the cortical precursors. During the migration of thymocytes to medulla, they maturated by well-defined selection processes (Fowlkes and Pardoll, 1989; von Boehmer, 1994). At the each maturation steps, they exhibit unique phenotypic features according to the expression of CD4 and CD8 coreceptor proteins and TCR receptors (Fowlkes and Pardoll, 1989). Generally immature thymocytes are CD4- CD8- (double negative) and CD4+ CD8+ (double positive), whereas in the medulla, they express CD4 or CD8 coreceptors and known as single positive (Nossal, 1994).
Apoptosis is a crucial mechanism for the maintenance of cellular homeostasis during development, differentiation and pathological processes. It also have a major function in the generation of lymphoid system and T cell development for the generating of T cell repertoire, deleting cells expressing unproductive TCR or exhibiting hypo-or hyper-responsiveness upon encountering MHC/self peptide complexes (Zacharchuk et al., 1991; Page et al., 1996). On the other hand, differentiation of most T lymphocytes is characterized not only by the exhibiting specific cell markers at each maturation stages, but also by changes in the glycosylation motifs.
In this study, it was investigated that, apoptotic thymocytes are reside in cortical regions of thymic tissues in the days 14 and 15 of embryonic development. After the 16th day, apoptosis was seen in cortex, corticomedullar junction and in medulla. The number of the apoptotic thymocytes in these regions was increased in 5 and 13 days old newborn mice. Fas molecule, a member of TNF-receptor superfamily that plays a unique role in apoptosis of lymphoid cells (Strasser, 1995; Nagata, 1997; Wallach et al., 1998), may participate in the normal apoptosis of thymocytes. Abundant expression of Fas was detected on CD4+CD8+ double positive as well as CD4+ or CD8+ single positive thymocytes in wild-type mice (Ogasawara et al., 1995).
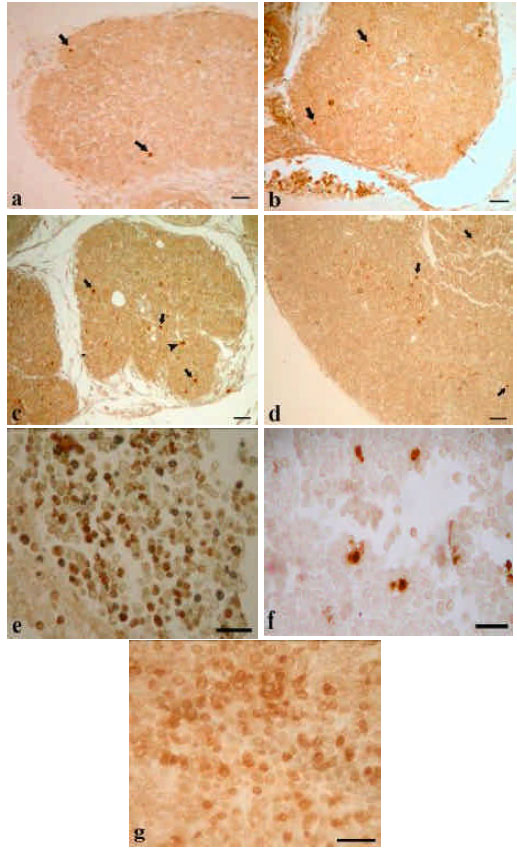 | |
| Fig. 1: | TUNEL stainings of embryonic and neonatal thymocytes. a-d represent embryonic thymocytes from 14, 15, 16 and 17th days of embryonic development, respectively; e and f, are thymi from 5 and 13th day after the birth, respectively. Arrows indicate cortical and medullar apoptotic cells of embryonic thymi. g, RNAse free DNAse treated control tissue. Bars 100 μm |
 | |
| Fig. 2: | PA-P-TCH-SP-PD stainings of embryonic thymi. Arrowhead indicates positively stained thymocytes, arrow points PA-P-TCH-SP-PD negative cells. Bar 10 μm |
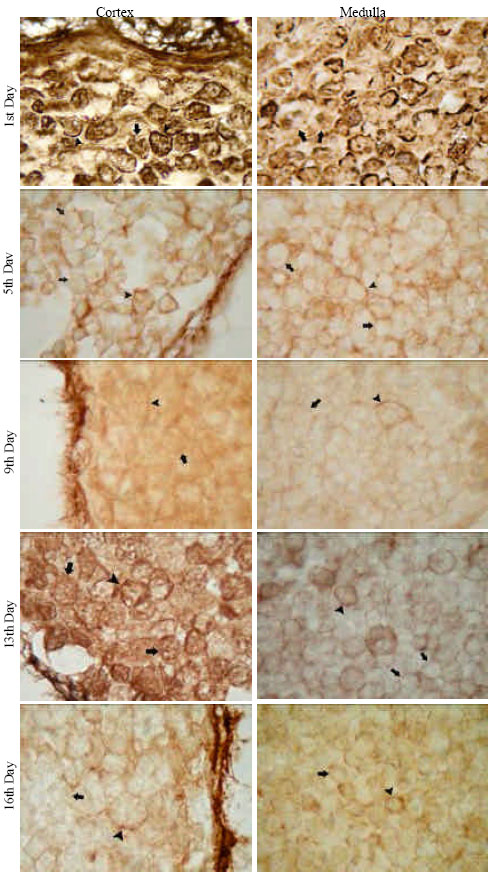 | |
| Fig. 3: | PA-P-TCH-SP-PD stainings of neonatal thymi. Arrowhead indicates positively stained thymocytes, arrow points PA-P-TCH-SP-PD negative cells. Bar 10 μm |
During the mouse development, from 16.5 day onward, Fas mRNA was detected in distinct cell types, including developing thymocytes (French et al., 1996). Interestingly, our TUNEL findings indicated that, apoptotic thymocyte number was increased after the 16th embryonic day. For this reason, during the late embryonic development, Fas induced apoptosis may be responsible for the deletion of thymocytes. On the other hand, although it was suggested that neonatal thymocytes do not express Fas (Miyawaki et al., 1992), number of apoptotic cells was increased in neonatal periods when compared the embryonic thymi (Fig. 1). In this instance, during the neonatal period, thymocyte apoptosis may be induced by another pathway such as galactin-1. In this regard, to evaluate the possible relationships between apoptosis and sialic acid expressions in embryonic and newborn thymi, we performed a histochemical staining method. During the thymocyte maturation, a number of reports demonstrated that the cell surface sialylations may change (Despont et al., 1975; Reisner et al., 1976; Akashi et al., 1988; Whiteheart et al., 1990; Alvarez et al., 1999). Our results indicated that there is a discrepancy in cell surface sialylation intensities between the embryonic and neonatal thymocytes (Fig. 2 and 3). They were abundant in embryonic thymi when compared to newborn ones. It is interestingly that, in the embryonic and neonatal periods, there is an inverse ratio between the increasing number of apoptosis with decreasing of sialic acid intensities. These investigation confirmed that the reduction in sialic acid presented on cell surface markers sensitizes a cell to apoptosis (Suzuki et al., 2003; Keppler et al., 1999).
During the T cell development, galactin-1 molecule induces T cell apoptosis (Perillo et al., 1997). Susceptibility of thymocytes to galactin-1 induced apoptosis may be related with specific oligosaccharides on the cell surface, because this molecule preferentially bound to thymocyte subsets that did not bind sambucus nigra agglutinin which recognize α2,6-linked sialic acids (Baum et al., 1995).
There are some differences between galactin-1 and Fas induced apoptosis. First, protein-protein interaction (i.e. Fas-FasL) is responsible for the Fas-induced apoptosis, whereas galactin-1 binds to cell surface glycoconjugates on thymocytes (Perillo et al., 1997). When the sialic acid level and apoptotic cell number differentiations considered in embryonic and newborn thymi, different pathways may responsible for thymocyte deletion in these two group. Consequently, during the late embryonic period (after the 16th day), due to masking effects of sialic acids, Fas-induced apoptosis may cause deletion of thymocytes. Nevertheless, after the birth, a reduction in sialic acids may give rise to galactin-1 mediated thymocyte apoptosis.
As a result, sialylation is emerging as an important issue in developing thymocytes and its intensity is related with pathways which responsible for thymocyte apoptosis.
ACKNOWLEDGMENTS
We thank Miss Pervin Toparlak for the elaboration of tissue sections. This work financed by Celal Bayar University Scientific Research Projects Committee (FEF 2002-103) and supported by Celal Bayar University Animal Ethic Council (Protocol No: 2004-70).
REFERENCES
- Ashwell, G. and J. Harford, 1982. Carbohydrate-specific receptors of the liver. Annu. Rev. Biochem., 51: 531-554.
CrossRefPubMedDirect Link - Cohen, J.J. and R.C. Duke, 1992. Apoptosis and programmed cell death in immunity. Ann. Rev. Immunol., 10: 267-293.
PubMed - Feizi, T., 1993. Oligosaccharides that mediate mammalian cell-cell adhesion. Curr. Opin. Struct. Biol., 3: 701-710.
CrossRefDirect Link - Keppler, O.T., M.E. Peter, S. Hinderlich, G. Moldenhauer and P. Stehling et al., 1999. Differential sialylation of cell surface glycoconjugates in a human B lymphoma cell line regulates susceptibility for CD95 (APO-1/Fas)-mediated apoptosis and for infection by a lymphotropic virus. Glycobiology, 9: 557-569.
Direct Link - Kerr, J.F., A.H. Wyllie and A.R. Currie, 1972. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer, 26: 239-257.
PubMedDirect Link - Manley, N.R. and C.C. Blackburn, 2003. A developmental look at thymus organogenesis: Where do the non-hematopoietic cells in the thymus come from?. Curr. Opin. Immunol., 15: 225-232.
CrossRefDirect Link - Schauer, R., 1982. Chemistry, metabolism and biological functions of sialic acids. Adv. Carbohydr. Chem. Biochem., 40: 131-234.
PubMed - Suzuki, O., Y. Nozawa and M. Abe, 2003. Sialic acids linked to glycoconjugates of Fas regulate the caspase-9-dependent and mitochondria-mediated pathway of Fas-induced apoptosis in Jurkat T cell lymphoma. Int. J. Oncol., 23: 769-774.
Direct Link - Varki, A., 1997. Sialic acids as ligands in recognition phenomena. FASEB J., 11: 248-255.
PubMedDirect Link - Wallach, D., A.V. Kovalenko, E.E. Varfolomeev and M.P. Boldin, 1998. Death-inducing functions of ligands of the tumor necrosis factor family: A Sanhedrin verdict. Curr. Opin. Immunol., 10: 279-288.
CrossRefDirect Link