
F. Azam
Nuclear Institute for Agriculture and Biology, P.O. Box 128, Jhang Road, Faisalabad, Pakistan
F. Aziz
Nuclear Institute for Agriculture and Biology, P.O. Box 128, Jhang Road, Faisalabad, Pakistan
S. Farooq
Nuclear Institute for Agriculture and Biology, P.O. Box 128, Jhang Road, Faisalabad, Pakistan
A. Lodhi
Nuclear Institute for Agriculture and Biology, P.O. Box 128, Jhang Road, Faisalabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2006 | Volume: 9 | Issue: 7 | Page No.: 1381-1388







ABSTRACT
A greenhouse experiment was conducted to assess the chamber effect on growth and nitrogen fixation in Sesbania aculeata (L.) at two levels each of soil salinity (electrical conductivity 4.65 and 7.23 dS m-1) and moisture (15 and 25%, v/w). The plants were grown either in the open or placed in an open-top polyethylene chamber and harvested 6 weeks after seed sowing. Salinization of soil led to a decrease, while higher moisture caused an increase, in different growth attributes of the plants. Growth reduction due to salt stress was less than that due to low moisture stress. Plants kept in the open-top polyethylene chamber showed better growth than those placed outside (in the open); all the parameters studied were better in the chamber. Chamber effect measured as Biomass Enhancement Ratio (BER) averaged 1.22 for different salinity and moisture treatments. However, root biomass of plants grown in the chamber was less than those grown in the open. Negative effect of salinity and low moisture was mitigated to a significant extent under chamber conditions. The analysis of root and shoot material for total N and 15N showed significant amounts of N2 being fixed as measured by isotopic dilution technique. Significantly higher amounts of N were fixed at 25 than 15% soil moisture; soil salinity had a depressing effect on the amount of N2 fixed. The contribution of biologically-fixed N (Ndfa) to the total N content of shoot and root portions was 23-62% and 21-52%, respectively, under different growth conditions. Contribution of Ndfa decreased with salinity but was more at 25% than 15% soil moisture.
PDF Abstract XML References Citation
How to cite this article
F. Azam, F. Aziz, S. Farooq and A. Lodhi, 2006. Chamber Effect on Growth and N2 Fixation of Sesbania aculeata (L.) At Two Salinity and Moisture Regimes. Pakistan Journal of Biological Sciences, 9: 1381-1388.
DOI: 10.3923/pjbs.2006.1381.1388
URL: https://scialert.net/abstract/?doi=pjbs.2006.1381.1388
DOI: 10.3923/pjbs.2006.1381.1388
URL: https://scialert.net/abstract/?doi=pjbs.2006.1381.1388
INTRODUCTION
Open top chambers have frequently been used to study the effect of elevated carbon dioxide (CO2) on plant growth, rhizodeposition and rhizospheric microflora and microbial functions. In these studies, enclosure of plants in the chambers is reported to have a significant positive effect (chamber effect) on growth and other attributes of ecosystem functioning (Sgheri et al., 1998; Mayr et al., 1999). Reduction in wind speed and turbulence relative to outside are considered to be the most important contributors to the chamber effect (Owensby et al., 1993). The resulting increase in humidity of the chamber will also lead to a reduction in evapo-transpiration and improved water use efficiency vis-à-vis stomatal responses. In addition, CO2 content of the chamber atmosphere relative to outside will certainly increase as a result of rhizorespiration (microbial and root respiration) and efflux of the resulting CO2. A substantial efflux of CO2 from soil is a common feature (Azam et al., 1985; Baggs et al., 2003; Pumpanen et al., 2003). This efflux will be sufficient to raise the CO2 concentration by several folds within the crop stand. The model proposed by Vaughan and Suarez (2002) shows CO2 efflux of ca 100 mg m-2 hr-1 from a wheat canopy. This efflux will be sufficient to raise the canopy CO2 level significantly with a substantial beneficial effect on plant performance (Drake and Leadley, 1991; Poorter, 1993; Idso and Idso, 1994; Azam and Farooq, 2001). The effect of CO2 has been more positive when environmental factors are affecting the plant growth and development severely (Koch and Monney, 1996).
Amongst different plant types, legumes occupy an important position with respect to the functioning of natural and artificial ecosystems through N acquisition from the atmosphere (Zanetti et al., 1997; Hopkins, 1999). In agroecosystems, green manuring legumes occupy a key position in maintaining/improving soil fertility and productivity. Their significance has increased further as a source of supplemental nitrogen due to economic and pollution potential concerns of chemical N fertilizers. Of the different legumes, species of Sesbania have generally been considered as most important for green manuring especially in wheat-rice rotation system. In arid regions, this system is characterized with varying degrees of salinity and water stress. However, there are hardly any studies to demonstrate chamber effect vis-à-vis elevated CO2 on green manuring legumes. Therefore, our objectives were I) to monitor changes in humidity, temperature, and CO2 outside and inside the chamber and ii) to study the response of Sesbania aculeata to different salinity and moisture regimes when grown under normal and changed (with respect to humidity, temperature and CO2) greenhouse conditions.
MATERIALS AND METHODS
Soil used in the experiment was collected from the top 0-15 cm of experimental plots with a history of receiving 15N-labelled fertilizer and cultivated to Sesbania. As such the soil was not only enriched with 15N (atom % 15N of the potentially mineralizable N being 0.3801 as determined by anaerobic incubation method (Stanford and Smith, 1972) but had sufficient number of rhizobia specific to Sesbania as suggested by dense nodulation in different experiments conducted previously. Air-dried and sieved (<2 mm) soil had the following characteristics: organic C, 0.6%; total N, 0.055%; NH4-N, 7 μg g-1 soil; NO3+NO2-N, 12 μg g-1 soil; pH (1:1, soil: water suspension), 7.7; electrical conductivity, 4.65 dS m-1; sand, 39.6%; silt, 26.2%; and clay, 34.2%.
Five-Kilogram portions of the air-dried and sieved soil were filled in 6-kg plastic pots and sown to Sesbania (5 seeds pot-1 on April 16, 2002); the seedlings were thinned to 3 within 2 days after seed germination. The soil was irrigated with water or water containing NaCl to achieve the variation in soil moisture content of 15 or 25% (v/w) and raise the salinity level to an electrical conductivity of 7.23 dS m-1. Nine pots were used for each treatment with triplicate being placed randomly in the I) greenhouse under ambient conditions and ii) open-top polyethylene chamber, and iii) open-top chamber to serve as control for irrigating the pots placed in (ii). The chamber consisted of an iron-bar frame (1 x 1 x 1.5 m) covered with polyethylene sheet and having an opening on the top to allow for gaseous exchange. Soil moisture content was maintained at required level by making up the weight loss determined in pots placed in one of the chambers used as control for this purpose. Loss of water was always less from enclosed than open pots. Relative humidity, temperature, and CO2 concentration inside and outside the chamber was monitored twice a day and 8 times during the experimental period of 6 weeks. Air samples obtained using 50-mL air-tight syringe were analyzed for CO2 on a gas chromatograph equipped with flame ionization and electron capture detectors.
Six weeks after seed sowing, data on number of leaves, maximum leaf length, and plant height were recorded. Uniform leaf samples were taken for the determination of Relative Water Content (RWC) of leaves by using the expression:
RWC = (fresh weight - dry weight)/(turgid weight - dry weight)*100
Turgid weight was determined by overnight incubation of leaf segments in water under dark conditions and blotting the extra water on leaf samples prior to weighing. The whole plants with intact roots were recovered from the pots by washing away the soil. All the nodules were carefully detached from the roots and weighed after drying at 65°C. Fresh and dry weight (oven drying a 65°C) of the root and shoot portions was recorded and the plant material finely powdered. Aliquots of the root and shoot material were analyzed for total N using micro-Kjeldahl method (Bremner, 1996). The distillates were acidified with 1M H2SO4 (0.1 μL μg-1 N), concentrated to 1 mL at 90°C on a hot plate, and appropriate volumes subjected to N-isotope analyses on a mass spectrometer using KOBr as alkalizer. Calculations for plant N derived from fixation were made on the basis of 15N dilution resulting from N2 fixation. Soil used in the present study was fairly enriched with 15N (atom % 15N content of potentially mineralizable N being 0.3801) thereby providing a reliable means of calculating the proportion of plant N derived from soil using the expression:
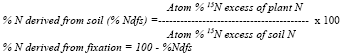 |
Amounts of plant N derived from soil and fixation were calculated by using the values obtained from above equations.
Statistical analyses of the data included determination of standard deviation of the means and coefficient of correlations using MS EXCEL computer software programme.
RESULTS
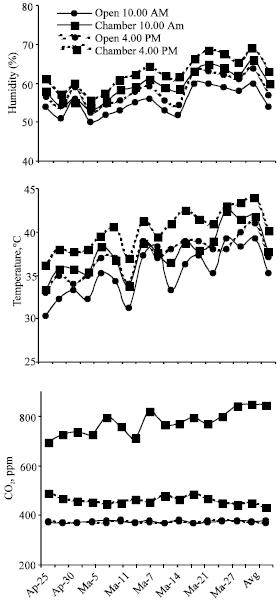
Temperature, relative humidity and CO2 were consistently higher in the chamber atmosphere as compared to that outside (Fig. 1). Observations recorded in the morning showed lower values for temperature and humidity but higher CO2 concentrations. In the present study, higher level of soil salinity had generally a depressing effect on the growth and biomass production of Sesbania (Fig. 2). All the growth parameters studied i.e., plant height, number of leaves, longest leaf, biomass of root and shoot, leaf water content, and nodule weight decreased due to salinity under all growth conditions. Relative water content (RWC) of leaves varied between 71 and 85% in different treatments. Lower values were obtained for plants grown in saline soil at lower (15%) moisture content. Under these conditions, a significant positive chamber effect was noted and RWC increased from 75 to 84% and from 71 to 85% at the two salinity levels. The differences were observed mainly at 15% moisture, while at 25% moisture, RWC was almost similar in different treatments. The results presented here suggest that salinity combined with low moisture exerted higher stress; while high moisture fairly mitigated the negative effects of salinity and there was no significant chamber effect either.
 | |
| Fig. 1: | Changes in humidity, temperature and CO2 concentration of the atmosphere in and outside the chamber during the months of April (Ap) and May (Ma). Average (Avg) of all the measurements are also plotted |
Salinity-induced reduction in growth was more pronounced at lower than that at higher soil moisture content. Growth reduction due to stress (GRS) (Poorter and Perez-Soba, 2001) was found to be 8.6%. Enclosure of plants in the open-top chamber had in general a positive effect (chamber effect) on different plant growth parameters especially under saline and low moisture conditions. Chamber effect expressed in terms of biomass enhancement ratio (BER) (Poorter and Perez-Soba, 2001) i.e., Biomass in the chamber/biomass in the open was 1.22 showing 22% gain in the biomass of plants grown under different conditions of salinity and moisture. Biomass of plants was significantly more at higher than that at lower soil moisture level (8.5 and 7.1 g pot-1, respectively, when expressed as an average of different treatments) with a negligible chamber effect in the former case; BER for higher and lower moisture regimes being 1.1 and 1.0, respectively. However, when lower moisture was taken as stress factor, GRS was 19%. Of the different plant parameters studied, nodulation as measured by biomass, showed a higher negative (to salinity) or positive (to moisture and enclosure in chamber) response. Chamber effect on nodulation in terms of BER was found to be 28%. Allocation of dry matter and N to roots was higher in plants grown in the chamber than those grown in the open i.e., 35 and 22% compared to 29 and 19%, respectively.
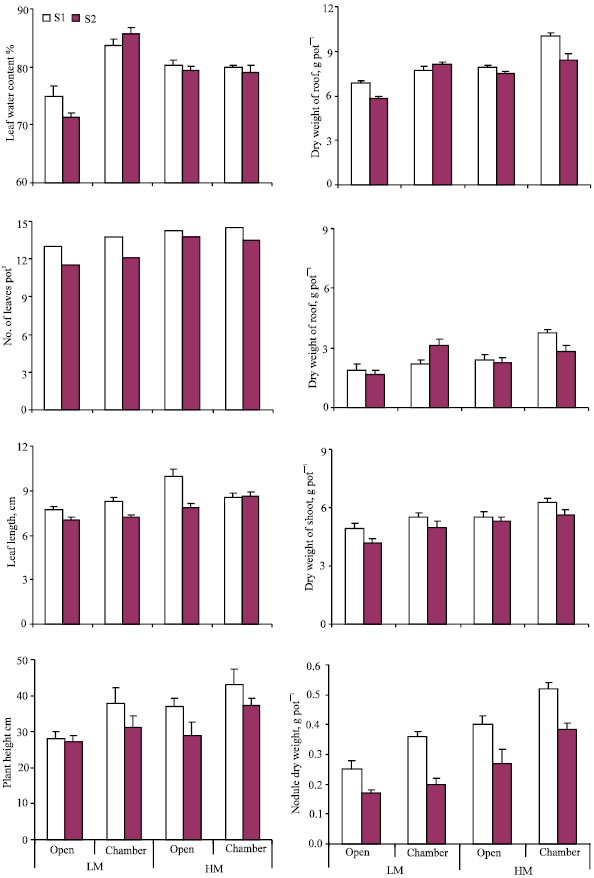
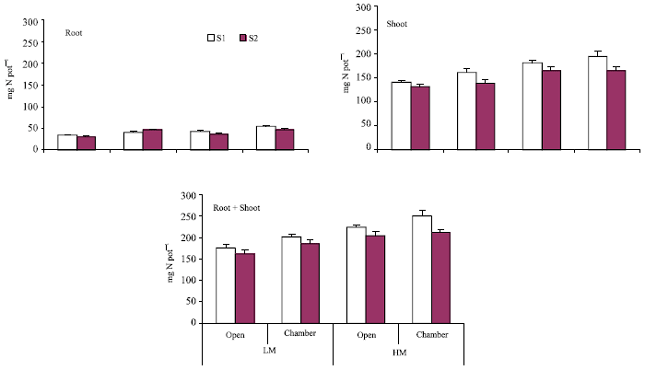
Trends in N content of root and shoot were similar to those observed for dry matter yield (Fig. 3). Computation of data in Fig. 3 and 4 showed a significant positive correlation of 0.77, 0.99, and 0.94 between dry matter and N content of shoot, root and shoot + root, respectively (n = 8 in each case). In the present study, roots were found to bear abundant pinkish nodules suggesting the presence of effective rhizobia in sufficient numbers. Hence, it was possible to determine the contribution of biological N2 fixation to the total N in plants using 15N isotopic dilution technique with some modification. In practice, a non-leguminous reference crop is required to estimate N2 fixation in the legume. In the present study, however, the difference in 15N abundance of soil N and atmospheric N was sufficiently high to enable the determination of percent N derived from soil as described in M & M section. The amount of N fixed varied between 36 and 147 mg pot-1 in different treatments (Fig. 4). Soil salinity had a significant depressing effect with the amount of N fixed being 79 mg pot-1 compared to 102 mg pot-1 under non-saline conditions. Higher moisture had a positive effect on the amount of N2 fixed with 122 and 59 mg N pot-1 being derived from fixation at 25% and 15% moisture, respectively. A greater proportion of fixed N was allocated to the shoot portion i.e., 77-84% of the total in different treatments. A positive chamber effect (averaging 41%) on the amount of N2 fixed under different conditions of salinity and moisture was observed.
Contribution of biologically fixed N to the total N content of shoot and root portions (%Ndfa) was 23-63% and 21-49%, respectively, under different growth conditions (Fig. 4). Lowest values were recorded for plants grown in the open at 15% soil moisture and highest values for those grown in the chamber at 25% moisture. On an average, the contribution of biologically fixed N remained low in non-saline soil. Overall contribution of fixed N to the total plant N ranged between 20-60%. A positive chamber effect of 30% was observed for the percent contribution of biologically fixed N to the total N of plants grown under two salinity and moisture regimes.
 | |
| Fig. 2: | Effect of salinity (S1, normal; S2, saline) and moisture (LM, 15%; HM, 25%) on different parameters of plants grown outside (open) and inside the chamber; standard error bars are also shown |
 | |
| Fig. 3: | Distribution of N in root and shoot portions of plants grown outside (open) and inside the chamber under different conditions of salinity (S1, normal; S2, saline) and moisture (LM, 15%; HM, 25%); standard error bars are also shown |
DISCUSSION
Atmospheric concentration of CO2 has increased significantly over the past few decades and is likely to double in the present century (King et al., 1992). The increase could benefit the plants in terms of increased photosynthesis and productivity (Poorter, 1993; Idso and Idso, 1994). The beneficial effects are generally more when plants are facing rhizospheric or atmospheric stresses (Koch and Monney, 1996; Polley et al., 1996). In most of such studies, the plants have been subjected to ambient or twice the ambient concentrations of CO2 using open-top chambers or free-air CO2 enrichment facilities. Results presented in Fig. 1 show that plants grown in open top chamber are in fact exposed to elevated levels of CO2, humidity and, temperature. Increase in CO2 concentration in the chamber can be attributed mainly to rhizorespiration (includes microbial and root respiration). In most studies, however, contribution of rhizorespiration to CO2 concentration within the plant canopies/stand has not been given due concentration. This happened in spite of the fact that, efflux of CO2 from soil is substantial and may potentially raise its level several times the ambient (Baggs et al., 2003; Vaughan and Surez, 2002).
Results presented in this study, show that mere enclosure of plants in an open-top chamber leads to improved growth and other plant functions, particularly the process of N2 fixation. Significant chamber effects have been reported by others (Sgheri et al., 1998; Mayr et al., 1999). In addition, stress imposed by salinity and low soil moisture content is mitigated to a significant extent by the conditions created in the chamber, especially a rise in relative humidity and CO2 concentration (Fig. 1). In the chamber, absence of free flow of wind (Owensby et al., 1993) could favour build-up of humidity leading to changes in evapotranspiration as well as photosynthesis vis-à-vis increase in the levels of CO2. Maintenance of higher soil moisture level in the enclosed pots was evident from less watering requirement (data not presented) and a higher RWC of leaves (Fig. 2). Besides high humidity, elevated CO2 is reported to decrease stomatal conductance by 30-60% (Morison, 1993) thus reducing water loss by plants. Therefore, elevated CO2 in the chamber atmosphere would have alleviated water stress leading to a substantial increase in N2 fixation and other growth attributes of Sesbania. Alfalfa (Medicago sativa) plants grown at elevated CO2 showed a lower decrease in leaf water potential than those grown at normal CO2 when subjected to water stress; chamber effect was noted (Sgheri et al., 1998). Similar results have been reported by others for different plant species (Rabha and Uprety, 1998; Seneweera et al., 2001).
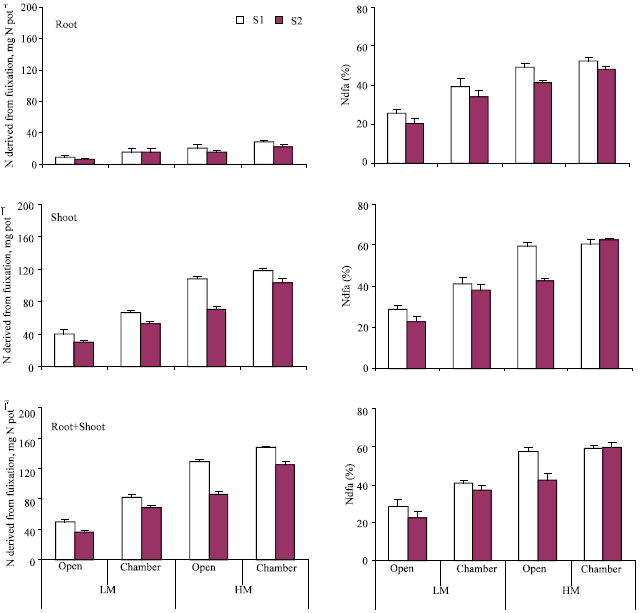 | |
| Fig. 4: | Contribution of biologically fixed N to the total N of root, shoot and root+shoot of plants grown outside (open) and inside the chamber under different conditions of salinity (S1, normal; S2, saline) and moisture (LM, 15%; HM, 25%); standard error bars are also shown |
In the present study, elevated CO2 would have mitigated the negative effects of increased temperature and humidity in the chamber. Higher moisture level in the root-zone had a positive effect on different growth parameters as well as N content of plants (Fig. 2 and 3). This could partially be attributed to enhanced N2 fixation under high moisture conditions that limit the availability of oxygen at the nitrogenase level; a condition suitable for efficient N2 fixation (Sangakkara et al., 1996). Since legumes depend mainly on biological N2 fixation besides N from soil organic matter, the factors that support the former process will have a positive impact on plant growth. Higher moisture level also mitigated the negative effect of salinity probably because of dilution of salts in the soil solution. This would also result in better plant growth especially at higher salinity level.
The observed increase in plant growth under chamber conditions could be attributed mainly to increased nodulation and enhanced N2 fixation (Fig. 4) at the expense of increased photosynthesis under elevated CO2 conditions. Stimulation of symbiotic N2 fixation at higher levels of CO2 (Yu et al., 2002) and a significant increase in different yield parameters of soybean have also been reported (Amthor et al., 1994). Partitioning of photosynthates to the roots, nodule mass, and rhizobial population are also reported to increase at elevated CO2 levels (Tschaplinski et al., 1993; Drake et al., 1997; Murillay et al., 1999).
Data presented in Fig. 4 shows 53-77% of the plant N being derived through biological nitrogen fixation and the rest (53-77%) from soil. This percentage is relatively lower than that commonly reported, i.e., 80-90% (Eaglesham et al., 1977) but possible as the plants were harvested at an early stage of growth. A higher contribution of biologically fixed N to the total plant N at elevated levels of CO2 is often attributed to increased C availability for nodule formation and for the functioning of nitrogenase (Tissue et al., 1997; Serraj et al., 1998). An increase in total N yield and the percentage of plant N derived from symbiotic N2 fixation in Trifolium repens under elevated CO2 (Zanetti et al., 1997).
Different factors had important bearing on the process of N2 fixation. Soil salinity had a negative effect while high moisture a substantially positive effect on the contribution of biological N2 fixation to the total plant N. This was attributable to the overall effect of these factors on plant growth. Reduced plant growth with increased salinity would lead to less photosynthesis and partitioning of photosynthates to the soil to support N2 fixation at higher salinity level. Enhanced rates of photosynthesis, on the other hand, should allow greater partitioning of carbohydrates below-ground, thereby enhancing root growth and enabling plants to better explore the soil for water and nutrients (Wall, 2001). In the present study, high moisture promoted the contribution of N2 fixation to N nutrition of plants. Contribution of BNF (biological nitrogen fixation) decreased with salinity, but elevated CO2 arrested the decrease to a significant extent. Such benefits are reported to be derived from the availability of more solutes for osmoregulation and from reduction in the transpirational intake of salts (Bazzaz, 1990). Munns et al. (1999) observed Salinity and CO2 interaction at low but not at high salinity have been reported (Munns et al., 1999). In addition, positive effect of elevated CO2 on root proliferation and root-induced microbiological and biochemical changes may help plants withstand salinity stress.
Results of this study suggest a positive effect of elevated CO2 on growth and N2 fixation of S. aculeata grown under mild salinity and high moisture. It is possible, therefore, to enhance the biomass yield of this green manuring crop by elevating the level of CO2 in the plant canopy. Under practical agricultural conditions, plant residues decomposing on the soil surface following mulching may help elevate the level of CO2 and thus the plant growth.
REFERENCES
- Azam, F., K. Haider and K.A. Malik, 1985. Transformation of 14C labelled plant components in soil in relation to immobilization-remineralization of N fertilizer. Plant Soil, 86: 15-25.
CrossRefPubMedDirect Link - Baggs, E.M., M. Richter, G. Cadisch and U.A. Hartwig, 2003. Denitrification in grass swards is increased under elevated atmospheric CO2. Soil Biol. Biochem., 35: 729-732.
CrossRef - Bremner, J.M., 1996. Nitrogen-Total. In: Methods of Soils Analysis, Part 3: Chemical Methods, Sparks, D.L. (Ed.). Soil Science Society of America, Madison, WI., USA., pp: 1085-1121.
Direct Link - Drake, B.G., M.A. Gonzalez-Meler and S.P. Long, 1997. More efficient plants: A consequence of a rising atmospheric CO2. Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 609-639.
Direct Link - Owensby, C.E., P.I. Coyne, J.M. Ham, L.A. Aven and A.K. Knapp, 1993. Biomass production in a tallgrass prairie ecosystem exposed to ambient and elevated CO2. Ecol. Appl., 3: 644-653.
Direct Link - Pumpanen, J., H. Ivesniemi and P. Hari, 2003. A process-based model for predicting soil carbon dioxide efflux and concentration. Soil Sci. Soc. Am. J., 67: 402-413.
Direct Link - Seneweera, S., O. Ghannoum and J.P. Conroy, 2001. Root and shoot factors contribute to the effect of drought on photosynthesis and growth of the C4 grass Panicum coloraum at elevated CO2 partial pressures. Aust. J. Plant Physiol., 28: 451-460.
Direct Link - Sgheri, C.L.M., M.F. Quartacci, M. Menconi, A. Raschi and F. Navari-Izzo, 1998. Interactions between drought and elevated CO2 on alfalfa plants. J. Plant Physiol., 152: 118-124.
CrossRefDirect Link - Stanford, G. and S.J. Smith, 1972. Nitrogen mineralization potentials of soils. Soil Sci. Soc. Am. J., 36: 465-472.
Direct Link - Tissue, D.T., J.P. Megonigal and R.B. Thomas, 1997. Nitrogenase activity and N2 fixation are stimulated by elevated CO2 in a tropical N2-fxixing tree. Oecologia, 109: 28-33.
Direct Link - Vaughan, P.J. and D.L. Suarez, 2002. Modeling above-canopy CO2 flux and evapotranspiration in wheat. Environ. Poll., 116: S37-S44.
CrossRef - Wall, G.W., 2001. Elevated atmospheric CO2 alleviates drought stress in wheat. Agric. Ecosyst. Environ., 87: 261-271.
CrossRefDirect Link - Yu, M., Q. Gao and M.J. Shaffer, 2002. Stimulating interactive effects of symbiotic nitrogen fixation, carbon dioxide elevation and climatic change on legume growth. J. Environ. Qual., 31: 634-641.
Direct Link - Zanetti, S., U.A. Hartwig, A. Luscher, T. Hebison and M. Frehner et al., 1997. Stimulation of symbiotic N2 fixation in Trifolium repens L. under elevated atmospheric CO2 concentration in grassland ecosystem. Plant Physiol., 112: 575-583.
Direct Link