
Shaha Ranajit Kumar
Department of Biochemistry and Molecular Biology, University of Rajshahi, Rajshahi-6205, Bangladesh
N. K. Sana
Department of Biochemistry and Molecular Biology, University of Rajshahi, Rajshahi-6205, Bangladesh
I. Hossain
Department of Biochemistry and Molecular Biology, University of Rajshahi, Rajshahi-6205, Bangladesh
Pakistan Journal of Biological Sciences
Year: 2004 | Volume: 7 | Issue: 1 | Page No.: 36-41







ABSTRACT
Seeds storage nutrient substances like protein, fat and carbohydrate used up during germination for seedling growth. Brassica seeds (Brassica napus L) were germinated at various concentrations of NaCl and Na-Phosphate buffer up to 200 mM. Germinating levels, seedling length, degradation of seed storage substances and lipase activity were investigated. High salt concentrations resulted in retardation of seed germination compared with buffer and distilled water treatment. Degradation of storage substances like free sugar, reducing sugar, starch, lipid and protein were severely reduced by elevated concentration of salts doses higher than 100 mM and Na-Phosphate buffer doses higher than 150 mM, respectively. The mobilization of storage lipid in water and buffer treated Brassica seedling started about 22 and 32 h respectably after imbibitions but salts treated germinating Brassica seedling delayed about 48 h after imbibitions. Lipase activity inhibited by salts treatment was confirmed by using triolein as a substrate.
PDF Abstract XML References Citation
How to cite this article
Shaha Ranajit Kumar, N. K. Sana and I. Hossain, 2004.
Ionic Effect on Mobilization of Seed Storage Nutrient Substances and Lipase Activity in Germinating Oil Seeds (Brassica napus L.). Pakistan Journal of Biological Sciences, 7: 36-41.
DOI: 10.3923/pjbs.2004.36.41
URL: https://scialert.net/abstract/?doi=pjbs.2004.36.41
INTRODUCTION
In plant seeds the storage nutrient substances are protein, fat and carbohydrate. During germination these storage nutrients used up for seedling growth (Harry, 1989). In plants accumulating a large proportion of carbon reserves as triacylglycerols (TAG), such as rapeseed (Brassica napus) and Arabidopsis, the activation of the β-oxidation and glyoxylate cycle during germination ensures conversion of fatty acid to carbohydrates necessary for the growth of the seedling before establishment of photosynthesis (Dieuaide et al., 1992; Pistelli et al., 1996). This is the first step of TAGs conversion to sugars required for growth of the germinating embryo (Ben Miled et al., 2000). Effect of various salts is a major environmental factor, which limits the growth and yield of crops around the world (Benziont et al., 1992). The effects of salinity have been studied at various stages of plant growth. It affects several aspects of plant metabolism, including lipid metabolism (Wu et al., 1998; Kuiper, 1985). The germination of lipid-rich seeds such as rapeseed has been to involve, among other processes, the rapid mobilization of storage triacylglycerols (TAGs) in the cotyledons of seedlings. Such hydrolysis of TAGs is catalyzed by highly active lipases. Enzymes involved in the process of lipid mobilization, such as malate synthase and isocitrate lyase, are detectable towards the late stages of embryo development (Chia et al., 2000). However, if much is known about the effect of salinity on crop plants, very few works have deals with the physiology of oilseeds (Gopal and Rao, 1982; Kayani et al., 1990). There have been very few reports on the effect of salts on degradation of seed storage substances particularly on oil seeds. Therefore, the aim of the present work was to study the influence of different doses of salts and buffer solutions on germination, seedling growth, degradation of seeds storage substance and lipase activity of Brassica napus L seeds during germination.
MATERIALS AND METHODS
Plant material: For experimental purpose oil seeds (Brassica napus L ) was collected from Bangladesh Agriculture Research Institute, Irshwdhi, Pabna and Rajshahi Local Shaheb Bazar Market. Good and mature seeds shorted out and was allowed to germinate in Petri dishes on filter paper moistened with distilled water containing sodium chloride (NaCl) salts and Na-Phosphate (Na2HPO4-NaH2PO4)-buffers at different concentrations ( 0, 50, 100, 150 and 200 mM ). Petri dishes were maintained in the light at 25°C. At different time interval (0, 24, 48, 72, 96 and 120 h), cotyledons were separated from seedlings, rinsed with distilled water and store at 4°C for biochemical analysis.
Biochemical analysis: Degradation of seeds storage substances sugar, protein and fats of Brassica seed during germination, treated with different concentrations of salts and buffers were determined by using standard procedures (Jayaraman, 1985; Wong, 1923; Lowry et al., 1951; Bligh et al., 1959; Morse, 1947; Bradford, 1976; Miller, 1959). Approximately 1.0 g of germinated seeds / Cotyledons (400mg) was ground in a mortar. The powder (pasty) was extensively extracted with methanol at 65°C in a Soxhlet apparatus for 2-3 h. Filtered over glass wool and was concentrated the oil materials using a Rotovapor (Flawil, Switzerland). The residual methanol extracted cake used for sugar and protein estimations during germination.
Preparation of extracts for lipase: Cotyledons were excised from seedlings (500-1000 mg) and then ground gently with a mortar and pestle in 5 ml of the pre-cooled grinding medium described by Murphy and Cummins (1989). The filtered homogenate was used as the crude lipase preparation. Sub cellular fractionation was obtained by differential centrifugation according to the method of Lin. et al. (1983). Homogenates prepared from cotyledons were centrifuged at 10000 g for 30 min at 4°C and the pellet was re-suspended in 1 ml of cold grinding medium. The 10000 g supernatant was further centrifuged at 100000 g for 2 h at 4°C and the resulting pellet was re-suspended in 1 ml of cold grinding medium and considered as microsomal fraction.
Enzyme assays: The lipase activities preparations were quantified by the hydrolysis of triolein (Macedo et al., 1997). One unit (U) of lipase activity was defined as 1 mmol of oleic acid released per minute at 37°C. Specific activity of lipase was expressed as the enzyme unit per mg of protein. All the experiments were carried out in triplicate.
RESULTS AND DISCUSSION
Brassica seeds germinated rapidly in distilled water, since after 22 h of imbibitions the germination level exceeded 65%. However, there was retardation in germination of salts-treated Brassica seeds. Germination started 24 h in the buffer treated seeds and 32h in the salts treated seeds after of imbibitions. On the third day (72 h) germination levels were higher than 80% for all applied buffer and salt doses upto150 mM (Fig. 1 and 2). In contrast, growth of seedlings as measured by length of redicals was severely depressed by salt concentrations higher than 100 mM and by buffer at 150 mM, respectively. For the latter concentration, germination and seedling growth were totally inhibited as compared with control (Fig. 3, 4). This finding is in good agreement with previous studies on sunflower (Ghasalli and Cherif, 1979), groundnut (Gopal and Rao, 1982) and Jojoba (Kayani et al., 1990) seeds germinating in saline conditions.
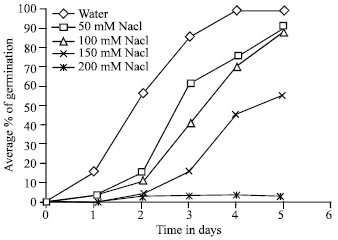 | |
| Fig. 1: | Effect of salt concentration during germination rate |
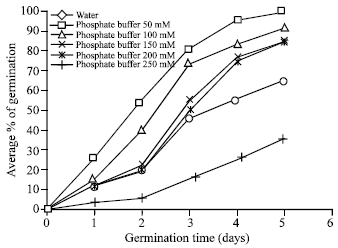 | |
| Fig. 2: | Effect of Phosphate buffer on germination of brasscia seeds |
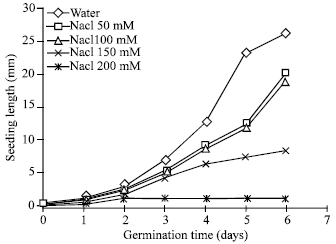 | |
| Fig. 3: | Effect of Nacl salt on seedling length of Brassica seeds during germination |
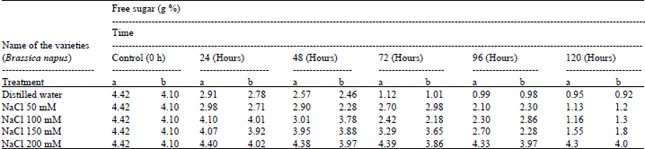
The free sugar and reducing sugar content in intact Brassica seeds were found to be 4.42 g%; 93.37 mg% and in the cake after oil extraction 4.10 g%; 90.10 mg%, respectively. During germination up to 120h free sugar and reducing sugar found regular degraded in intact Brassica seeds 0.95 g% and 12.2 mg% (Table 1, 2).
| Table 1: | Free sugar content of Brassica seeds using different ionic media during germination |
 | |
| Table 2: | Reducing sugar content of Brassica seeds using different ionic media during germination |
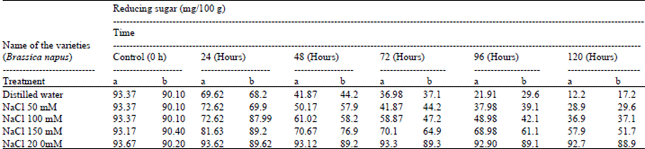 | |
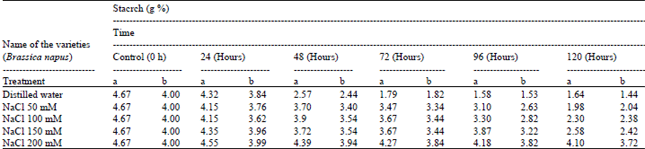
| Table 3: | Starch content of Brassica seeds using different ionic media during germination: |
 | |
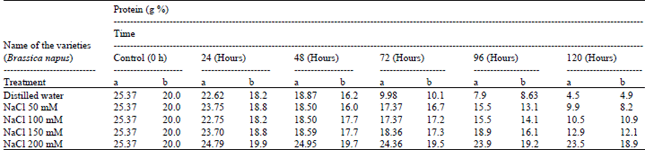
| Table 4: | Total protein content of Brassica seeds using different ionic media during germination |
 | |
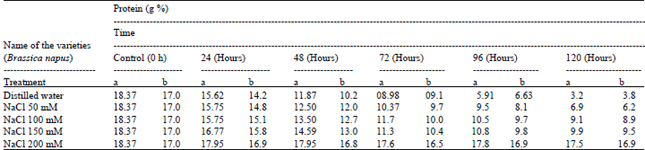
| Table 5: | Water-soluble protein content of Brassica seeds using different ionic media during germination |
 | |
| a: Intact seeds powder b: Cake after oil extraction | |
| Table 6: | Effect of various NaCl concentrations on TAG degradation of Brassica cotyledons. (Results are expressed in mg of TAG / 10 cotyledon pairs) [Different concentration of NaCl solutions] |
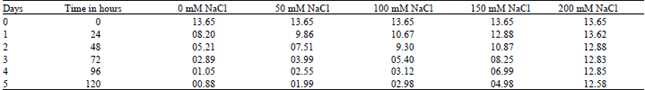 | |
| Table 7: | Effect of various Na-Phosphate Buffer (Na2HPO4 - NaH2PO4 concentrations on TAG degradation of Brassica cotyledons. (Results are expressed in mg of TAG / 10 cotyledon pairs) [Various concentration of Na-P- buffer] |
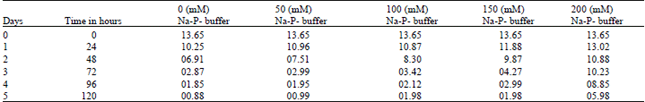 | |
| Table 8: | Effect of various NaCl doses on lipase activity from sub cellular fractions of Brassica seeds cotyledons (72h). ((Activity is expressed in ηmol of 17.5/min per cotyledon pair) |
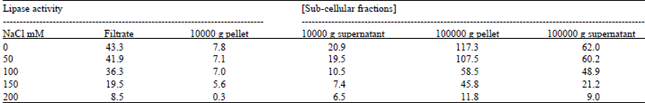 | |
| Table 9: | Effect of various Na-Phosphate (Na2HPO4 - NaH2PO4 buffer doses at pH: 6.9 on lipase activity from sub cellular fractions of Brassica cotyledons(72h). (Activity is expressed in ηmol of 17.5/min per cotyledon pair) [sub-cellular fractions] |
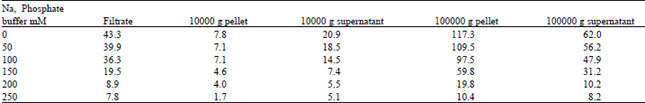 | |
But degradation of free sugar is inhabited due to salts and buffer solutions (about 40%). One of the nutritional reservoirs in plants, which used as a fuel, is starch. Starch was found to be 4.67 g% in intact seeds, which degrade to 1.64% upto120h. The present findings clearly revealed that the composition of starch in Brassica seeds degraded up to 48h of germination then constant during further germination in control and in different concentrations of salts doses (Table 3). In light germination probably starch firstly degrade for embryo growth then other enzyme utilize seed storage substance for the energy supply for seedling growth.
Total protein and water-soluble protein contents were 25.37 and 18.37 g% in the intact seeds, which degrade to 4.5 and 3.2 g% during germination upto120h. Protein degradation of the germinated Brassica seeds at different doses of salts were shown in Table 4 and 5. The present results clearly demonstrated that the percentage of protein (water soluble and insoluble) present in Brassica seeds enhanced mobilization during further germination (72-120 h) in presence of different concentrations of salts and buffer solution. This indicates that proteolytic enzyme also inhibited by salts and buffer solutions.
Treatment with salts higher than 100 mM and buffer150 mM inhibit germination as a results degradation of nutrient also inhibited. The degradation of nutrient components namely sugar, protein and starch were inhibited not much and the results are in agreement with earlier studies by Gad et al., (1965). Degradation data sugar, protein and starch by buffer solution are not shown.
Oil component in the Brassica seeds degraded sharply during germination due to the high lipase activity, which was also inhibited by salts and buffer treatments.
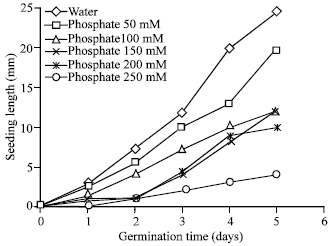 | |
| Fig. 4: | Effect of Phosphate buffer on seedling growth |
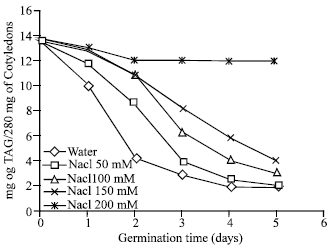 | |
| Fig. 5: | Effect of Nacl solution on TAG mobilization during germination of Brassica seeds |
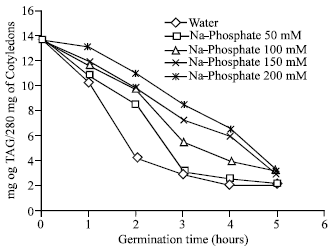 | |
| Fig. 6: | Effect of Na-Phosphate buffer on TAG mobilization during germination of Brassica seeds |
(Table 6, 7). In oil seeds remarkable changes occur in oil degradation during germination, which was studied earlier by (Miled Ben et al., 2000).
As shown in Table 6 and 7, germination of Brassica seeds was accompanied by a considerable mobilization of triacylglycerols (TAG). TAG contains the major form of lipid reserves, representing 92% of total lipids. In control cotyledons, TAG molecules were quickly hydrolyzed, since by the 2nd (Second) day of germination more than the half of initial TAG content was broken down and by the 4-5th day the remaining TAG represents only 2.5% of initial amount, where as in salts and buffer treatments cotyledons, there was a delay in degradation that increased with salts (100 mM) and buffer (150 mM) concentrations in the growing medium. The delay in TAG degradation caused by salts and buffer stress is probably due to an inhibition of lipase involved in TAG hydrolysis in cotyledons of germinated Brassica seeds.
In vivo lipase activity, from cotyledons of Brassica seedling treated with different salts concentrations was investigated to enzyme extracts of 3-day old cotyledons. The highest lipase activity was observed in control extracts when about 75% of TAG was hydrolyzed (Fig. 5 and 6). This TAG degradation was sharply inhibited in NaCl salts solution and slightly phosphate buffer-treated extracts, especially at elevated concentrations. Thus the seedlings subjected to 150 mM salt solution, this activity was totally inhibited and TAG levels were equal to those of dry seeds (Table 6and 7). Sub cellular fractionation of total extract from control, salts and buffer treated cotyledons showed that the most lipase activity occurred in the microsomal fraction (100000 g pellet), as has been reported in previous studies (Hills et al., 1988; Murphy and Cummins, 1989). Lipase activities from sub cellular fractions (Table 8 and 9) were found to decrease under increasing NaCl salts and phosphate buffer concentrations, which confirmed our above results on crude lipase activity.
In conclusion, the decrease in seedling growth caused by salts and buffer treatment may be correlated with inhibition of TAG hydrolysis, a process that generates sugars for the growth of the germinating embryo. Furthermore, inhibition of lipase in cotyledons from salts and buffer treated seedlings could be explained by a direct effect of Na+, Cl–, PO43–ions on the enzyme, reducing its activity or by an indirect effect of salts by modifying gene expression of the enzyme and then affecting its de novo synthesis. However, it is well known that within the cells ions are compartmentalized, which may exclude the direct effect of salts and buffer.
ACKNOWLEDGEMENT
Author of this paper grateful to Dr. Rezaul Karim, Department of Biochemistry and Molecular Biology, University of Rajshahi for his help for preparing this manuscript.
REFERENCES
- Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Macedo, G. and G.M. Pastore, 1997. Lipases microbianas na producao de esters formadores de aroma. Ciencia Tecnol. Alimentos, 17: 115-119.
Direct Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Morse, E.E., 1947. Anthrone in estimating low concentration of Sucrose. Anal. Chem., 19: 1012-1013.
Direct Link