
Parichehr Hanachi
Department of Biochemistry and Microbiology, Faculty of Science and Environment Studies, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
N. A. Shamaan
Department of Biochemistry and Microbiology, Faculty of Science and Environment Studies, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
J. Ramli
Department of Biochemistry and Microbiology, Faculty of Science and Environment Studies, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
J. H. Arshad
Department of Biochemistry and Microbiology, Faculty of Science and Environment Studies, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M. A. Syed
Department of Biochemistry and Microbiology, Faculty of Science and Environment Studies, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2004 | Volume: 7 | Issue: 2 | Page No.: 209-211







ABSTRACT
The glutathione S-transferase (GST) were affinity purified in both mice treated with 200 mg kg-1 benzo (a) pyrene once weekly for 8 weeks and control mice. The affinity purified enzymes were isoelectric focused and three zones were selected in each group of mice. GST in the BaP treated mice were focused into zone I (pI 7.5 ), zone II (pI 8.4) and zone III (pI 9.4). In control mice, the zones were zone I (pI 8.4), zone II (pI 9.2 ) and zone III (pI 9.8. The affinity purified and focused enzymes were subjected to SDS-PAGE and assayed for substrate specificities. The SDS-PAGE of the affinity purified GST revealed 3 subunits of molecular radius of approx. 23, 24.5 and 26 kD in control mice and 24, 26.5 and 28 kD in BaP treated mice. The activities of the focused GST towards 1-chloro-2,4-dinitrobenzene (CDNB) were similar in both BaP treated and control mice. Activities of focused enzymes in the BaP treated mice towards 3,4-dichloronitrobenzene (DCNB) and ethacrynic acid (EA) were higher than control mice. The findings suggested that the GST in BaP treated mice were different to that in control mice. The SDS-PAGE showed that the subunits in the BaP treated mice were larger than control mice.
PDF Abstract XML References Citation
How to cite this article
Parichehr Hanachi, N. A. Shamaan, J. Ramli, J. H. Arshad and M. A. Syed, 2004. Evidence That Benzo (a) Pyrene Induced Large Glutathione S-Transferase Subunits in the Mouse Mus musculus. Pakistan Journal of Biological Sciences, 7: 209-211.
DOI: 10.3923/pjbs.2004.209.211
URL: https://scialert.net/abstract/?doi=pjbs.2004.209.211
DOI: 10.3923/pjbs.2004.209.211
URL: https://scialert.net/abstract/?doi=pjbs.2004.209.211
INTRODUCTION
The glutathione S-transferase (GST) are dimeric enzymes involved in dedetoxication of foreign compounds. In particular, the over expression of GST in tumours appear to be a factor in the development of acquired resistance towards anticancer drugs and hence GST are a therapeutic target for rational drug design[1].The hepatic GST appear to play an important role in reducing the BaP-induced DNA damage in target and non-target tissues[2].
The dimmers can consist of homodimers or heterodimers[3]. The dimmers have been classified according to the pI values, molecular size, specificities to different substrates, amino acid sequence and immunochemical properties. The monomers ranged in size between 19-27 kD and normally distinguished by SDS-PAGE. The expression of the GST have been reported to alter after exposure to various compounds, notably to carcinogens e.g. benzo (a) pyrene (BaP). Induction of GST by the carcinogens may be due to the overexpression of individual/several existing monomers or new monomers resulting in increased enzyme activity. We had purified GST by affinity chromatography in the liver of mice treated with the carcinogen benzo (a) pyrene and control mice, partially separated the different forms of the affinity purified GST by isoelectrofocusing (IEF) and characterized the GST of the benzo (a) pyrene treated mice in comparison with control mice.
MATERIALS AND METHODS
Chemicals glutathione: 1-chloro-2,4-dinitrobenzene and benzo (a) pyrene, immobilized glutathione on agarose matrix and all other reagents used were of highest grade commercially available (Sigma Chemical Co., St. Louis, Mo, USA).
Treatment of animals: The mice Mus musculus were divided into two groups of control and treated. Treated group were injected once a week interaperitoneally with 200 mg kg-1 BaP dissolved in corn oil while one group was injected once a week interaperitoneally with corn oil and acted as control.
Preparation of cytosol: The mice were then left for 8 weeks before being sacrificed by cervical dislocation. Immediately after sacrifice the liver was excised and the extraction of GST initiated. Fresh mice livers were weight and were rinsed in KCl 1.15% buffer.
Enzyme assays: The cytosolic fraction were prepared using the method of Speier and Wattenberg[4]. GST assay was determined according to method of Habig et al.[5] and protein determination was carried out according to the method of Bradford[6].
GST purification: The ammonium sulphate precipitation 40-80% and affinity chromatography on agarose matrix was used for purification of GST. Active fraction from affinity chromatography were pooled and isoelectric focusing electrophoresis using the rotofor cell (Bio Rad, USA) was carried out toward CDNB, DCNB and EA as substrate. SDS-PAGE and NATIVE-PAGE were carried out according to the method of Laemmli[7].
RESULTS
The affinity purified GST were partially separated by isoelectric focusing into three forms into three forms of enzymes with different pI values; benzo (a) pyrene treated mice-pI 7.5, 8.4 and 9.4; control mice-pI 8.4, 9.2, 9.8 (Fig. 1).
The enzymes in each pI zones were then subjected to SDS-page and molecular size of the subunits estimated by regression analysis. The subunit molecular weight were estimated to be 23, 24.5 and 26 kD in control mice, 24, 26.5 and 28 kD in the BaP treated mice (Fig. 2).
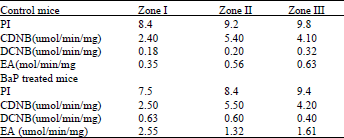
The enzymes in each pI zones were also assayed for activities towards different substrates (Table 1). Generally, the GST in the BaP treated mice had significantly higher (p<0.05) activities towards the substrates than those in the control.
DISCUSSION
The GST obtained in the BaP treated mice had different characteristics with regards to pI values, subunit size and substrate activities compared with control mice. BaP treatment over a long period induced different GST in the mice. One of the GST subunit in the BaP treated mice was larger than that in control mice and not found in control mice. The pI values were different in the BaP treated mice and these enzymes had different activities towards the substrates used.
Purified GST in the mouse have been reported to be homodimeric and had subunits of approx. 24, 25.5 and 27 kDa, corresponding to the designation of MII, MI and MIII, respectively[8]. In contrast, our findings showed that the affinity purified GST to be heterodimers showed that the affinity purified GST to be hetrodimers with subunits approx. The GST in treated sample exhibits electrophoretic migration on SDS- PAGE closely similar to cytosolic GST in control.
Estimation of subunit size yielded varied results due to several factors including the use of different types of protein standards, methods for separating the isoforms and the conditions of the experiments[8,9]. Mouse GST were found to be homodimeric by choromato focusing[8] but we obtained heterodimers by isoelectric focusing. Probably, IEF may be less able to completely separate the various GST forms, resulting in a mixture of the various forms.
GST in the BaP treated mice had higher activities towards DCNB and EA than those in control mice. The activities towards CDNB were similar in both the mice since CDNB is a universal substrate for the GST. The higher activities towards DCNB and EA in the BaP treated mice are different to the control mice. Mouse GST MII similar and in the same class as rat GST 7-7 and human GST-π, had higher activities towards DCNB and EA than mouse GST MI and MIII[10].
Warholm et al.[11] had described one isoform of GST in the human liver, which had a high affinity for BaP oxide.
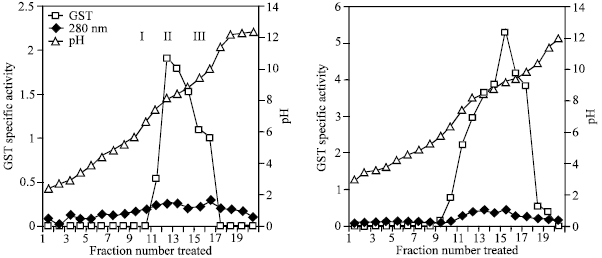 | |
| Fig. 1: | Isoelectric focusing of affinity purified GST in Benzo (a) pyrene treated and control mice |
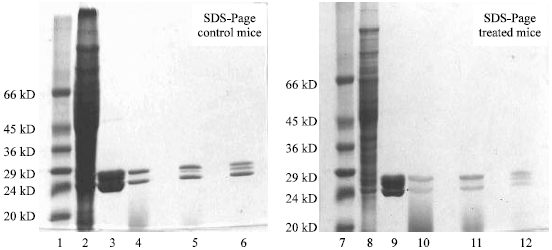 | |
| Fig. 2: | SDS-PAGE of affinity purified and isoelectrofocused GST fractions 1, protein standard; 2, Cytosolic fraction; 3, Affinity purified fraction; 4, pI 8.4; 5, pI 9.2; 6, pI 9.8; 7, protein standard; 8, cytosolic fraction; 9, Affinity purified; 10, pI 7.5; 11, pI 8.4; 12, pI 9 |
| Table 1: | Activities of GSTs in IEF zones towards different substrates.The isoelectrofocused enzymes were assayed for activities towards 1-chloro-2,4-dinitrobenzene (CDNB), 3,4-dichloronitrobenzene (DCNB) and ethacrynic acid (EA) |
 | |
It could be possible that one of the GST forms detected in this study could act on BaP oxide but no attempt was made to determine the activity of the affinity purified GST towards BaP oxide.
The affinity purified GST in the BaP treated mice were different from that in control mice as shown by subunit size, pI values and activities to the different substrates. The peaks were present as different GST components that were responsible for transcriptional activation of GST genes maybe increases in levels of relevant mRNA.
ACKNOWLEDGMENT
The authors would like to acknowledge Universiti Putra Malaysia and IRPA for supporting this study.
REFERENCES
- Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link