
Maliha Rashid
National Agriculture Research Center, Park Road, Islamabad, Pakistan
Samina Khalil
National Agriculture Research Center, Park Road, Islamabad, Pakistan
Najma Ayub
National Agriculture Research Center, Park Road, Islamabad, Pakistan
Sadia Alam
National Agriculture Research Center, Park Road, Islamabad, Pakistan
Farooq Latif
National Agriculture Research Center, Park Road, Islamabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2004 | Volume: 7 | Issue: 2 | Page No.: 187-196







ABSTRACT
After screening a large number of phosphate solubilizing microorganisms (PSM) isolated from rice rhizosphere on the basis of solubilization index (SI) and pH drop ten best bacterial strains (10RB, 54RB, 57RB, 20RB, 21RB, 23RB, 22RB, 13RB, 20RB, 40RB) and three best fungal strains {(Aspergillus flavus(1RF), Penicillium canescens(4RF), A. niger (8RF)}were further analyzed for their acid production ability using HPLC technique. SI of 13 best isolates ranged from 2.16-6.23. SI and change of pH of these isolates showed abrupt changes during seven days of incubation in Pikovskayas agar and broth medium respectively. This drop of pH clearly indicated the production of organic acids. HPLC results confirmed that few PSM isolates produced gluconic, fumaric, succinic, acetic and some unknown organic acids in smaller concentrations while oxalic and citric acids in larger concentrations in their broth medium. Among bacterial strains 10RB produced largest amount of organic acids (2.4747 g-l) while among fungal isolates Aspergillus flavus produced greatest amount of acids (1.835 g-l) than the other two fungal strain. The concentration of glucose when analyzed during HPLC was found less in all the cultural samples as compared to the control (13 g-l) that was without any PSM culture. This shows that all the PSM strains utilized the carbon of glucose for the production of organic acids. The abilities of these efficient strains for phosphorus (P) solubilization and immobilization were also quantitatively investigated in Pikovskayas broth medium. The results showed that all the strains solubilized (0.04-0.1%) and immobilized (0.1-0.4%) P but fungi immobilized more P than bacteria. Correlation among all the parameters showed a positive(r = 0.2, P<0.1) correlation between the organic acid excretion and P solubilization and a negative correlation was found between pH and P solubilization (r =-0.4, P<0.1) and immobilization (r=-0.5, P<0.05).
PDF Abstract XML References Citation
How to cite this article
Maliha Rashid, Samina Khalil, Najma Ayub, Sadia Alam and Farooq Latif, 2004. Organic Acids Production and Phosphate Solubilization by Phosphate Solubilizing
Microorganisms (PSM) Under in vitro Conditions. Pakistan Journal of Biological Sciences, 7: 187-196.
DOI: 10.3923/pjbs.2004.187.196
URL: https://scialert.net/abstract/?doi=pjbs.2004.187.196
DOI: 10.3923/pjbs.2004.187.196
URL: https://scialert.net/abstract/?doi=pjbs.2004.187.196
INTRODUCTION
Microorganisms are important component of soil. Soil bacteria and fungi mediate soil processes such as decomposition, nutrient mobilization and mineralization, storage release of nutrients and water, nitrogen fixation and denitrification. In the frame of agriculture, the microflora is of great significance because it has both beneficial and detrimental influence upon man’s ability to feed himself (Gaur, 1990; Motsara et al., 1995; White law, 2000).
PSM include different types of microorganisms that convert insoluble phosphatic compounds into soluble forms (Prerena-Akhaury et al., 1997; Raju and Reddy, 1999). Important genera of phosphate solubilizing bacteria are Bacillus and Pseudomonas (lllmer and Schinner, 1992; Motsara et al., 1995). Aspergillus and Penicillium are important P solubilizing fungal genera (Motsara et al., 1995). It has been reported that certain strains of Rhizobium can also solubilize both organic and inorganic phosphates (Abd-Alla, 1994). Isolated acid and alkaline phosphate producing bacteria and fungi from 13 different soil types and vegetation. It has been observed by many investigations that a high proportion of P solubilizing microorganisms are concentrated in the rhizosphere of plants (Gaur, 1990). Since phosphate activities are found to be much higher in rhizosphere soil than in bulk soil (Seeling and Jungk, 1992). Inorganic phosphorus solubilizing microorganisms are more concentrated in rhizosphere plants than in bulk soil (Vesquez et al., 2000).
Many researchers have quantitatively investigated the ability of PSM to solubilize insoluble P in pure liquid medium cultures. A wide range of liquid media supplied with nutrients that are required for the growth of PSM and with different P sources has been used in order to study the nutritional effects on the growth and physiological properties of PSM strains. The levels of P solubilized by PSM and metabolites such as organic acids produced by PSM were also investigated in liquid cultures (Whitelaw, 2000; Narula et al., 2000).
Kucey et al. (1989) and Bar-yosef (1991) have shown that microbial solubilization of soil phosphates in liquid medium studies has often been due to the excretion of organic acids. In many studies the presence of organic acids i.e. oxalic acid, citric acid, lactic acid, gluconic acid etc. in liquid culture filtrates were determined by paper chromatography or thin layer chromatography and by modern techniques such as High Performance Liquid Chromatography (HPLC), lsotachophoresis and enzymatic methods have been used by others to allow more accurate identification of unknown organic acids. (Banik and Dey, 1982; 1983; Parks et al., 1990; Berthelin et al., 1991; Cunningham and Kuiack, 1992; Illmer and Schinner, 1992; 1995a; Gupta et al., 1994; Singal et al., 1994; Illmer et al., 1995; Vassilev et al., 1995,1996; Mehta et al., 1996; White law et al., 1999). Banik and Dey (1883) and Asea et al. (1988) detected organic acids in culture solutions of PSM but did not show any correlation between the solubilization of P and amount of organic acids produced by PSM.
Many research scientists have examined the reason that when phosphorus-solubilizing microbes were applied to the insoluble P, a larger amount of unavailable P became available (Barber, 1984; Subba Rao, 1984). The role of organic acids produced by PSM in solubilizing insoluble P may be due to the lowering of pH, chelation of cations and by competing with P for adsorption sites in the soil. It has also been investigated that organic acids may also form soluble complexes with metal ions associated with insoluble P (Ca, Al, Fe) and thus P is released (Kepert et al., 1979; Omar, 1997).
Some of the physiological properties of PSM that were investigated are nutritional requirements, oxygen–demands and temperature effects. Glucose is the main carbon source for the growth of PSM but other carbon sources can also be utilized (Illmer and Schinner, 1992; Motsara et al., 1995).
Inorganic anions also solubilize phosphorus but they are less effective than organic anions Kim et al. (1997a) reported that HCl was able to solubilize less P from hydroxyapatite than citric acid or oxalic acid at same pH.
The objective of this research was to evaluate the biochemical and physiological characteristics of indigenous rice rhizosphere PSM and to develop a better understanding of the accurate mechanism of P solubilization by correlating all the determined parameters.
MATERIALS AND METHODS
Solubilization index on solid and change in pH in PSM broth cultures: Sterilized PKV media was poured into sterilized petri plates after solidification of the media, a pinpoint inoculation of bacterial or fungal strains was made on the plates under aseptic conditions. The plates were incubated at 28±1°C for 7 days and observed for colony diameter and diameter of solubilization zone regularly during 7 days. Solubilization index was evaluated according to the ratio of the total diameter (colony + halo zone) and the colony diameter (Edi-Premono et al., 1996).
Change in pH by 10 PSB and 3 PSF in broth cultures was determined with the help of pH meter daily during seven days of incubation.
Phosphorus solubilizing capacity of PSM in liquid cultures: Phosphorus solubilizing capacity of 10 PSB and 3 PSF was determined in 100 ml PKV broth medium (Pikovskaya 1948). 0.25 g l-1 insoluble phosphate (in the form of tricalcium phosphate) was added to 100ml broth before sterilization then 0.5 ml suspension of each microbial culture was added in duplication to the broth after its sterilization. A control without any PSM was also maintained. The organisms were allowed to grow for five days at 30°C and then were individually centrifuged at 15000 rpm for 30 min. The supernatant was collected in 100 ml volumetric flasks and volume was made up to 100 ml with distilled water. While in case of fungi the centrifuged solution was filtered through What man filter paper No. 42 and the clear solution was collected in 100 ml volumetric flasks and the volume was made up to 100 ml with distilled water. Thus extract of each solution was prepared (Sultanpour and worksman, 1979). Then available phosphorus in the broth cultures was determined by Watenabe and Olsen (1965).
Immobilized phosphorus of microbial biomass in broth: For immobilized P of microbial biomass PSM were grown in PKV broth medium. Five days old cultures were individually centrifuged at 2000 rpm for 20 min. Microbial biomass after centrifugation in the pellet form was collected which was then digested by wet ash method (Chapman and Pratt, 1961). In this method microbial biomass was taken in Pyrex Kjeldahl’s flask and 10 ml mixed acid (perchloric and nitric acid) was added in each flask. Then flasks were heated on a hot plate at a temperature of 100-300°C until the solution samples (about 2-3 ml) became colorless. Flasks was removed from hot plate and cooled. Then each solution was transferred into a 50 ml volumetric flask and volume (50 ml) was made with distilled water. For P estimation, 5-ml extract of each sample was taken in a test tube and 5 ml color developing reagent was added. The tube was stirred in a mechanical stirrer. The reading was taken on spectrophotometer at 430 nm and the concentration of total immobilized P percentage in microbial biomass was calculated.
Organic acids produced by PSM with HPLC: For organic acid determination PSM were grown and multiplied in PKV liquid medium. On 5th day cultures were blended and were centrifuged at 2000 rpm for 20 min. Supernatant of each blended culture was filtered through 0.45 μm non sterile 4 mm sized micro filter syringes.
The 20 μl purified solutions of each culture was injected in a bio-rad ion exchange column of Aminex 87-H (25*4.6 mm) .The operating conditions consisted of 0.001 N H2SO4 the mobile phase at a constant (isochratic) flow rate of 0.6 ml min-1 and column was operated at 25°C. Organic acid concentrations in samples were determined with the help of RI detector.
The software used during HPLC was a Turbochrome navigator system. RI impulse was read with the help of Turbochrome navigator programmer in g l-1. The unknown organic acids in purified solution were determined by comparing the retention times and peak areas of chromatograms with the standards of oxalic acid, citric acid, gluconic acid, fumaric acid and acetic acid.
Statistical analysis: Correlation values among different parameter of the present study were determined by simple correlation coefficient regression equation. The result means were depicted diagrammatically by computer program MSTATC microsoft exel version 5.0 graphic facility.
RESULTS AND DISCUSSION
Biochemical and Physiological Studies of PSM: Solubilization Index on solid and change in pH in broth medium: Solubilization index (SI) of 10 PSB and 3 PSF strains was ranged from 2.16 to 6.23 in the present work (Fig. 1a, 2a). Sometimes abrupt changes occurred in these values similar observations have been reported by several workers (Subba Rao, 1984; Motsara et al., 1995; Kim et al., 1997b; Kumar and Narula, 1999; Gaur, 1990;White Law, 2000).
pH of bacterial broth dropped significantly (5.96 to 2.75) as compared to control where it remained constant at 7.1 (Fig. 2a). Three fungal strains also caused decrease in pH from 7.28(control) in the beginning to 4.5 within seven days (Fig. 2b). The pH drop in PSM liquid cultures have been reported in several researches which supports the pH change in present study (Cunningham and Kuiack, 1992; Motsara et al., 1995; Illmer et al., 1995; Bar Yosef et al., 1999).
Acid production in broth culture: HPLC of 13 efficient PSM (10 bacteria and 3 fungi) strains revealed that all of the isolates produced organic acids (Table 1).
Results of all the strains indicate that oxalic and citric acids were two major acids produced by all the tested PSM strains. Other organic acids produced by these strains were gluconic, succinic, fumaric and acetic acids. Among bacterial strains 10RB produced largest amount of acids than all other bacterial strains followed by 54RB and 13RB.
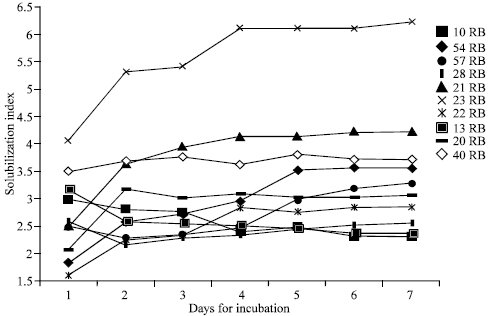 | |
| Fig. 1a: | Solubilization index of 10 efficient PSB strains during seven days of incubation |
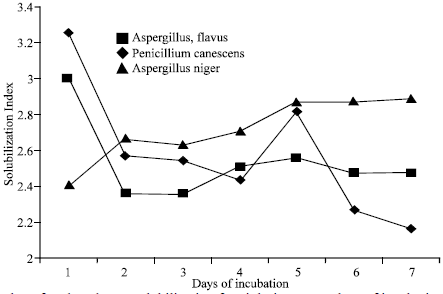 | |
| Fig. 1b: | Solubilization Index of 3 Phosphorus solubilization fungi during seven days of incubation Each value is mean of two replicates, Solubilization Index= Colony+halazone diameter/Colony diameter |
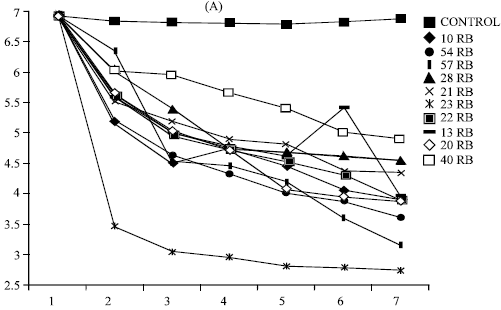 | |
| Fig. 2a: | Change in pH by 10 efficient phosphorus solubilizing bacteria strain in liquid Pikovskaya’s medium during 7 days of incubation Each value is mean of two replicates, RB= Rice Bacteria |
The fungal strain 1RF (A. flavus) produced acids in greater amount than the other two fungal strains of 4RF (P. canescens) and 8RF (A. niger). In case of A. niger oxalic acid and citric acid were two major acids. (Cunningham and Kuiack, 1992; Illmer et al., 1995) also reported the similar results. A. flavus and P. canescens in the present study produced oxalic acid, citric acid, gluconic acid and an additional succinic acid in case of A. flavus. Results in the present study are supported by the observation of Banik and Dey, (1982), (1983); Venkateswarlu et al. (1984); Parks et al. (1990); Gupta et al. (1994); Singal et al. (1994), White law et al. (1999) who studied the production of oxalic acid, citric acid, gluconic acid, succinic acid and tartaric acid etc. by Aspergillus and other fungal genera.
The result of A. niger contradict the result of Venkateswarlu et al. (1984) who reported that only lactic acid was produced by A. niger and to Vesquez et al. (2000) who reported that only succinic acid was produced by A. niger. This contradiction can be satisfied by giving the argument that strains used in the present study were different because these were isolated from different soils as Nautiyal et al. (2000) isolated stress tolerant PSM from alkaline soils and those were different from the non stress tolerant strains isolated from other soils.
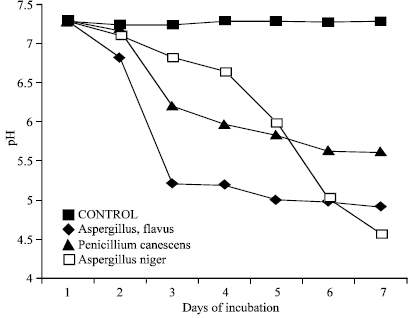 | |
| Fig. 2b: | Change in pH by 3 efficient phosphorus solubilizing fungal strain in liquid Pikovskaya’s medium during 7 days of incubation Each value is mean of two replicates |
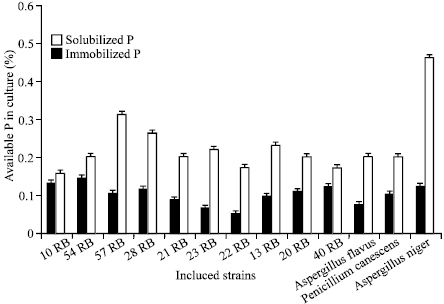 | |
| Fig. 3: | Solubilization and immobilization of tricalcium phosphate in five days old broth culture of efficient PSM strains Each value is the mean of two replicates, RB=Rice Bacteria, |
During HPLC of control without PSM, the glucose concentration was 13 g l-1. Glucose acted as a main carbon source for acid production in present study, therefore in all the cultures of inoculated PSM strains during HPLC unused glucose concentration was in the range of 0.1-10 g l-1. This result is supported by Gaur (1990) who reported that PSM are heterotrophs and solubilize insoluble phosphates by secreting organic acids, the role of carbon sources in this context is very important.
During present study Pikovskaya medium was modified by changing the composition with the addition of 13 g l-1 glucose instead of 10 g l-1. It is similar to the findings of Gaur (1990) who reported that by increasing the concentration of glucose from one to three percent in the medium, the role and the quantity of rock phosphate solubilized by phosphorus solubilizing fungal strains increased and also greater acidity was produced by glucose at the higher concentration, which is an important factor for phosphate dissolution.
The concentration of total acids, known acids and unknown acids along with pH drop caused by them in their broth culture are given in Table 1.
| Table 1: | Organic acid production and changed pH in five days incubated broth culture of 13 selected PSM strains |
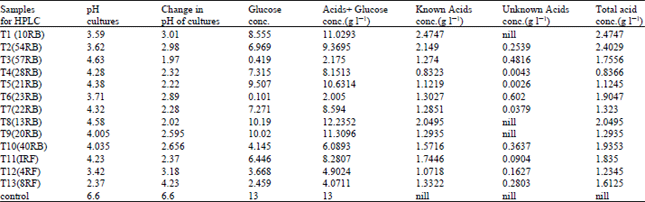 | |
| RB = Rice Bacteria, RF = Rice Fungi, Conc. = Concentration | |
| Table 2: | Correlation coefficients between different parameters of selected PSM strains studied on solid and liquid Pikovskaya’s medium under sterilized conditions |
 | |
| * significant, ** highly significant | |
Tricalcium phosphate solubilization by PSM in broth: P solubilized values in each broth sample shows that different strains solubilized the P at different rates. (Fig. 3) Phosphate solubilization of the added TCP in broth was in the range of 0.04 – 0.14%. Among individual cultures of all the bacterial strains 54RB and 10RB solubilized highest amounts of insoluble phosphorus i.e. 0.1417% and 0.1318% respectively. While among the fungal strains A. niger solubilized highest amount of supplied TCP. Among bacterial strains 57RB immobilized highest amount of available phosphorus (0.31%) while in case of fungal strains A. niger (8RF) immobilized highest amount (0.45%) of available phosphorus in culture. Overall fungal strains immobilize greater amounts of phosphorus than bacterial strains (Fig. 3). This is similar to the findings of Khalil (1995) that fungi immobilize more P than bacteria. The P content in the biomass of the PSMs was 0.1 to 0.4% for ten bacterial and three fungal cultures in the present study. Thus it’s confirmed that phosphorus like nitrogen is mineralized as well as immobilized. Phosphorus is essential for microbial cell synthesis. Fungi assimilate 0.24 to 0.4 parts of phosphorus for every 100 parts of glucose oxidized and Streptomyces sp assimilate 0.27 to 0.63 and 0.16 to 0.36 parts of phosphorus respectively in same conditions (Gaur, 1990).
Illmer and Schinner (1992) reported that P content in the biomass of PSMs were 0.4- 0.8% for Pseudomonas sp. and P. aurantiogriseum respectively, which is within the range given by Beever and Burns (1980) for ordinary microorganisms. They stated that very effective P uptake systems of microorganisms would enable the assimilation of P from the solution, by disturbing the equilibrium between P salts and P in solution this observation is supported by Illmer and Schinner (1995b) they observed highly significant correlation between P- mobilization and the production of microbial biomass. Jurinak et al. (1986) also gave the statement that solubilization mechanisms depend on the production of microbial biomass.
Correlation between different in vitro studied parameters: Data analysis gave a significant (r = -0.6, P≤0.01) correlation between pH and colony + halazone formation of 13 efficient PSM strains (Table 2).
pH and solubilization index were negatively correlated with correlation coefficient (r ) of –0.6. Negative correlation (r = -0.4, P≤0.1) between pH and solubilization of calcium phosphate minerals was found in the present study. pH of the liquid medium of 13 efficient PSM strains was negatively correlated with P solubilization (r = -0.4, P≤0.1). pH was also negatively correlated with the P immobilization (r = -0.5 at P≤ 0.05) . Significantly higher and negative (r= -0.6, P≤0.01) correlation was observed between the P solubilized on solid medium observed by halazone formation and pH drop in broth. This observation is supported by the findings of Venkateswarlu et al. (1984) where a significantly higher correlation (r = -0.93) between pH and P solubilization was found.
These findings were opposite to the results of Illmer and Schinner (1992), with correlation coefficient (r) value of 0.49. Illmer and Schinner (1992) reported that sometimes the culture filtrate pH was relatively high and yet in the medium high P solubilization occurred, this may occur due to the chelation of organic acids with Ca++ ion in tricalcium phosphate.
Similarly it has been reported by Asea et al. (1988), Salih et al. (1989) Whitelaw et al. (1999) that pH had no affect on P solubilization and found positive correlation between the pH and P solubilization. Narsian et al. (1995) also found no correlation between the pH and solubilization of P over an incubation period of 7 days. Similar observation was reported with P.aurantiogriseum by Illmer and Schinner (1992), with a Penicillium isolate by Goenadi and Saraswati (1993) and with P. radicum by white law et al. (1999).
Several authors attribute solubilization of inorganic P sources to the production and release of organic acids (mainly citric acid, oxalic acid and succinic acid). However, others suggested that organic acids may play an important role, but are not the only possibility for solubilizing RP, which is confirmed by the weak or poor correlation between pH and the amount of solubilized P (Asea et al., 1988; Ehrlich, 1990; Salih et al., 1989).
During present study positive but insignificant correlation between the organic acid excretion and P solubilization was observed.
Similarly Banik and Dey (1983) also detected organic acids in culture solutions but were not able to find a correlation between solubilized P on one hand and the amounts of organic acids produced by PSMs on the other hand. Therefore, reasonable doubts arose as to whether organic acids are really and exclusively involved in solubilization mechanisms (Banik and Dey, 1982; Asea et al., 1988). Illmer et al. (1995) observed that organic acid production may be helpful but not the sole need for AlPO4 solubilization.
It has been experimented that Pseudomonas sp. and Penicillium aurantiogriseum were very effective in solubilizing calcium phosphate without producing appreciable amounts of organic acids while observed that Aspergillus and Penicillium solubilized P by producing large amounts of organic acids. The production of citrate, oxalate and gluconate by Aspergillus niger has also been reported (Illmer et al., 1995).
It is also observed by Taha et al. (1969), Illmer and Schinner (1995a) that there is no correlation between P solubilization and organic acid excretion, the P solubilization was due to the release of protons accompanying respiration or NH4+ assimilation.
Opposite to the present findings the correlation between the organic acids excretion and P solubilization was significant as given by Nahas (1996), Vassilev et al. (1996) and White law et al. (1999) with r values of 0.57, 0.83 and 0.91 respectively indicating that acid production was the key P solubilization mechanism in many cases.
Solubilization index was also significantly correlated (r = -0.4, P≤0.1) with the P solubilized by 13 efficient PSM strains. The correlation between the colony diameter and halazone + colony diameter was positive and significant(r = 0.5, P≤0.05). The correlation (r= -0.4, P≤0.1) between the SI on agar medium and P solubilized in liquid cultures of 13 PSM strains is supported by the findings of Gupta et al. (1994) that some isolates with little clear zone on solid medium exhibited high efficiency for dissolving insoluble phosphates in liquid medium and some showed large clearance zones on agar but low P solubilization in liquid medium.
This shows that the plate technique is insufficient to detect all P solubilized as commented by Nautiyal (1999) that soil microbes should be screened in broth cultures for the identification of most efficient solubilizers. A significant correlation (r=0.70) between the ability of 23 fungal isolates to produce clear zone on precipitated phosphate agar and their ability to solubilize rock phosphate in liquid medium. Conversely, Cunningham and Kuiack (1992) reported that the day on which the clear zone is measured is important. It was observed in this study, that certain bacteria lost their P solubilization ability after sub culturing; Similarly high percentage of the bacterial isolates loose their abilities when sub-cultured.
It is concluded from the present study that all the isolated PSM strains produce organic acids by utilizing the carbon of given substrate in their broth medium. These PSM strains are also capable to solubilize and immobilize inorganic phosphorus provided in the broth medium. Further research should be conducted to evaluate the P solubilization efficiency of these PSM strains during in vivo studies.
REFERENCES
- Abd-Alla, M.H., 1994. Phosphatases and the utilization of organic phosphorus by Rhizobium leguminosarum biovar viceae. Lett. Applied Microbiol., 18: 294-296.
CrossRefDirect Link - Asea, P.E.A., R.M.N. Kucey and J.W.B. Stewart, 1988. Inorganic phosphate solubilization by two Penicillium species in solution culture and soil. Biol. Biochem., 20: 459-464.
CrossRef - Banik, S. and B.K. Dey, 1982. Available phosphate content of an alluvial soil as influenced by inoculation of some isolated phosphate-solubilizing micro-organisms. Plant Soil, 69: 353-364.
CrossRefDirect Link - Banik, S. and B.K. Dey, 1983. Alluvial soil microorganisms capable of utilizing insoluble aluminium phosphate as a sole source of phosphorus. Zentralblatt fur Mikrobiologie, 138: 437-442.
CrossRefDirect Link - Bar-Yosef, B., R.D. Rogers, J.H. Wolfram and E. Richman, 1999. Pseudomonas cepacia-mediated rock phosphate solubilization in kaolinite and montmorillonite suspensions. Soil Sci. Soc. Am. J, 63: 1703-1708.
CrossRefDirect Link - Beever, R.E. and D.J.W. Burns, 1980. Phosphorus uptake, storage and utilization by fungi. Adv. Bot. Res., 8: 127-219.
CrossRefDirect Link - Cunningham, J.E. and C. Kuiack, 1992. Production of citric and oxalic acids and solubilization of calcium phosphate by Penicillium bilaii. Applied Environ. Microbiol., 58: 1451-1458.
PubMedDirect Link - Gupta, R., R. Singal, A. Shankar, R.C. Kuhad and R.K. Saxena, 1994. A modified plate assay for screening phosphate solubilizing microorganisms. J. Gen. Applied Microbiol., 40: 255-260.
Direct Link - Illmer, P. and F. Schinner, 1992. Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Biol. Biochem., 24: 389-395.
CrossRefDirect Link - Illmer, P. and F. Schinner, 1995. Phosphate solubilizing microorganisms under non-sterile conditions. Bodenkultur, 46: 197-204.
Direct Link - Illmer, P. and F. Schinner, 1995. Solubilization of inorganic calcium phosphates-Solubilization mechanisms. Soil Biol. Biochem., 27: 257-263.
CrossRefDirect Link - Kim, K.Y., G.A. McDonald and D. Jordan, 1997. Solubilization of hydroxyapatite by Enterobacter agglomerans and cloned Escherichia coli in culture medium. Biol. Fertil. Soils, 24: 347-352.
CrossRef - Kim, K.Y., D. Jordan and G.A. McDonald, 1997. Effect of phosphate-solubilizing bacteria and vesicular-arbuscular mycorrhizae on tomato growth and soil microbial activity. Biol. Fertil. Soils, 26: 79-87.
CrossRefDirect Link - Nautiyal, C.S., 1999. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett., 170: 265-270.
CrossRefPubMedDirect Link - Nautiyal, C.S., S. Bhadauria, P. Kumar, H. Lal, R. Mondal and D. Verma, 2000. Stress induced phosphate solubilization in bacteria isolated from alkaline soils. FEMS Microbiol. Lett., 182: 291-296.
CrossRefPubMedDirect Link - Omar, S.A., 1997. The role of rock-phosphate-solubilizing fungi and Vesicular-Arbusular-Mycorrhiza (VAM) in growth of wheat plants fertilized with rock phosphate. World J. Microbiol. Biotechnol., 14: 211-218.
CrossRefDirect Link - Vassilev, N., M.T. Baca, M. Vassileva, I. Franco and R. Azcon, 1995. Rock phosphate solubilization by Aspergillus niger grown on sugar-beet wastemedium. Applied Microbiol. Biotechnol., 44: 546-549.
CrossRefDirect Link - Vazquez, P., G. Holguin, M.E. Puente, A. Lopez-Cortes and Y. Bashan, 2000. Phosphate solubilizing microorganisms associated with the rhizosphere of mangroves in a semi arid coastal lagoon. Biol. Fertil. Soils, 30: 460-468.
Direct Link - Watanabe, F.S. and S.R. Olsen, 1965. Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci. Soc. Am. J., 29: 677-678.
CrossRefDirect Link - Whitelaw, M.A., T.J. Harden and K.R. Helyar, 1999. Phosphate solubilisation in solution culture by the soil fungus Penicillium radicum. Soil Biol. Biochem., 32: 655-665.
CrossRefDirect Link - Khalil, S., 1995. Direct application of phosphate rock and appropriate technology fertilizers in Pakistan. Proceedings of the International Workshop on Direct Application of Rock Phosphate and Appropriate Technology Fertilizers in Asia-What Hinders Acceptance and Growth, Feb. 20-25, Kandy, Sri Lanka, pp: 231-236.
Mohammad Mosharraf Hossain Reply
Very good and effective paper for citation and further research indication path.