
M. Idrish Miah
Department of Physics, Bangladesh Institute of Technology, Chittagong, Chittagong-4349, Bangladesh
Anders Johnsson
Department of Physics, Norwegian University of Science and Technology, Trondheim, Norway
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 5 | Page No.: 478-484







ABSTRACT
Cell type studied in this work is human leukemic cell Jurkat. In order to identify the exponential growth phase, the better time for sampling the population, the cell line was evaluated by monitoring the proliferation and viability of the cells in a 7-days long culture adding no medium to the cells. It was found that the culture within the cellular concentration from 6x105-2.7x106 cells/ml was in the exponential or log phase, where the viability and reproduction for the cells were high. The lag time and population doubling time were calculated and found to be 1 and 5 days respectively. The density dependent survival of cells in culture was remained constant, with the exception at very higher cell densities where it was found to decrease slightly. The viability of the cells was found to remain within 96-97% at incubation temperature range 10-500C, with a maximum at 35-400C. The cells were then incubated with 1 mM ALA at different temperatures and irradiated with different light doses. The dose-response curves were studied and modeled. The cell survival for incubation temperatures 10-370C were modeled as sigmoidal Boltzmann curves, whereas the survival for temperatures 39-500C were modeled as the second-order exponential decay curves. The efficiency for cell inactivation was calculated as the light dose or exposure time required for 50% of cell survival, and it was found that the efficiency was higher at higher temperatures.
PDF Abstract XML References Citation
How to cite this article
M. Idrish Miah and Anders Johnsson, 2003. Study of Cell Line and Dose-response of Human Leukemic Cells. Pakistan Journal of Biological Sciences, 6: 478-484.
DOI: 10.3923/pjbs.2003.478.484
URL: https://scialert.net/abstract/?doi=pjbs.2003.478.484
DOI: 10.3923/pjbs.2003.478.484
URL: https://scialert.net/abstract/?doi=pjbs.2003.478.484
INTRODUCTION
Photodynamic therapy (PDT) has become a wide-speed technique in the treatment of cancer and other diseases that acts mainly via singlet oxygen (1O2) generated by energy transfer from photosensitizers excited with visible light (Dahle et al., 2000 and Kennedy et al., 1990). 1O2 has a very short lifetime in cells, so the intracellular targets are close to the sites where the sensitizer is localized. The subcellular localization of a sensitizer varies with its molecular characteristics such as lipophilicity, electrical charge and size as well as with the incubation time and washing technique (Berg et al., 1990).
Aminolevulinic acid (ALA) is the precursor of protoporphyrin IX (PPIX) in the heme biosynthesis pathway and has been suggested for photodiagnosis and PDT of cancer and other diseases because PPIX can function as photosensitizer (Miah, 2001a; Wyld et al., 1997; Dailey, 1990 and Bogorad, 1979). There are two rate-limiting steps in this process: synthesis of ALA from succinyl CoA and glycine by the ALA synthetase and conversion of PPIX to heme by the enzyme ferrochelatase (i.e., by inserting an iron atom into the porphyrin ring) (Hilf et al., 1999; Dailey, 1990 and Bogorad, 1979). Provision of cells with excess exogenous ALA bypasses the first rate-limiting step causing accumulation of PPIX that can be used in PDT. On exposure to light, reactive oxygen species are produced by energy transfer from excited triplet-state PPIX, causing cellular damage and
ultimately cell death (Hilf et al, 1999). The mode of cell death induced by PDT is dependent on the sensitizer, the cell line used and the cell density (Hilf et al., 1999). The damage may be direct or may result from hypoxia induced by shutdown of the tissue microcirculation. Microcirculatory shutdown has been shown to occur following PDT in vivo with both traditional porphyrin photosensitizers (Reed et al., 1989) and, recently, ALA-induced PDT (Roberts et al., 1994 and Leveckis et al., 1995). Because the rate of cell doubling may vary for different cell types the proliferetion of the cell line should be studied. The exponential or log phase of a culture is the best time for sampling the population in PDT because it is quite uniform and viability is high. In present work, cell line of human leukemic cell Jurkat and the ALA-induced dose-response curves of the cells have been studied. The main objective of this work was to find out an efficient temperature for PDT treatment of the cells.
MATERIALS AND METHODS
This experimental work was carried out at the Department of Physics, Norwegian University of Science and Technology, Norway during May, 2000 to March, 2001.
Chemicals: ALA and trypan blue were purchased from Sigma Chemical Co (Sigma-Aldrich, USA). A stock δ-Aminolevulinic acid (ALA) solution of 100 mM was prepared. The RPMI 1640 medium and L-glutamine were from Merck, USA, and fetal calf serum (FCS) was from Hgclone, USA. A 70% ethanol solution was used to wash the Bόrker Chamber.
Cell line and culture conditions: The Jurkat cell line (clone E6.1) purchased from the American Type Culture Collection (ATCC), USA was cultured in an incubator (Biomed CO2-incubator, ASSAB Classic) under 37°C temperature, 80% relative humidity and 5% CO2 conditions and maintained in a logarithmic phase (Fig. 1). The medium RPMI 1640 was supplemented with fetal calf serum (FCS) (10%), L-glutamine (2 mM) and gentamycine (40 μg ml–1). For keeping the cells in exponential phase within the cellular concentration of 105-106 cells ml–1 (for more detail, see growth curve in Fig. 1), the cell suspension was diluted every 3rd or 4th day during the culture. In all experiments, the cells were treated under sterile conditions. Much attention was given on CO2 supply and water circulation into the incubator to ensure the culture environment in the incubator.
Cell viability: A 100 μl-sample of cell suspension was mixed properly with 50 μl of 0.1% trypan blue (in 0.81% sodium chloride and 0.06% potassium phosphate solution). Trypan blue is excluded from viable cells, whereas non-viable cells with impaired membrane integrity take up the dye and appear blue on light microscopy (Jacob et al., 1985). Two drops of the resultant solution was placed on the plate of the Bürker Counting Chamber, and the cells, both live and necrotic, on the plate were counted by using a microscope with sufficient magnification.
Cell proliferation assay: The cell proliferation was studied by counting the cells in a continuous cell line culture every 24th hour for 7 days, without diluting the suspension in the flasks. The initial plating cell density was 3.7x104 cells ml–1. In order to calculate the doubling time (tD) and lag time for the cell type cell counting was performed on three flasks per day until a plateau was reached.
Irradiation and incubation: Cells proliferating in the exponential phase with cell density 1-2 x 106 cells ml–1 were incubated with 1 mM ALA for 4 hours and irradiated at room temperature (20± 2°C) using the irradiation system reported elsewhere (Miah, 2001b), with a fluence rate of 4.1 mW/cm2. The irradiance value at the cell position was about 2.5 mW/cm2. The emission was mainly in the wavelength region 400-500 nm, with a peak at 430 nm. After irradiation cells were again incubated for 4 hours. The same procedure was repeated in incubation at different temperatures. Cell survival was then studied by measuring the colony-forming ability of the cells.
Statistical analyses and modeling: All temperature experiments were performed several times (n ≥ 3) for error analysis. The survival curves for different temperatures were modeled by using the Origin 5.0 Professional program.
RESULTS AND DISCUSSION
The kinetics and proliferation of cells demonstrate that the rate of cell doubling may vary for different cell types (Griffiths, 1979). For this reason, the proliferation of the chosen cell line (Jurkat E6.1) should be studied first before dealing with the cells in any property-related investigations. The log phase of a culture is a period of exponential increase in cell numbers and is the most reproducible aspect of the culture, and it is the optimal time for sampling the population because it is quite uniform and the viability is high. It is proposed (Griffiths, 1979) that the viability of the cells during sampling should not be less than 95%. In order to maintain the culture in desired logarithmic phase, the rate of cell division and viability should be controlled. This can be done by administering to the cells, e.g., by controlling the amount (and times of adding) of the culture medium, under the defined culture protocol for the cell type.
The proliferation and viability is shown for the Jurkat E6.1 cell line during 7-day long continuous culture adding no medium to the cells (Fig. 1). The initial plating or seeding cell concentration was taken 3.7x104 cells ml–1. As seen in the figure, the viability during the first 6 days was remained constant at about 97% and then it was decreasing. The cell survival at the 7th day of the culture was less than 95%. From the proliferation curve in the figure, it can be seen that for the cell concentrations larger than about 2.7x106 cells ml–1 the slope of the curve might decrease, indicating that the culture was entered the plateau phase. This suggests that the cellular concentration for the chosen cell type should not exceed 2.7x106cells/ml. The log phase, as indicated by the two broken lines in Fig. 1, is the study phase in which the cells should be kept to maintain the culture in a uniform cell line with a high viability and a reproduction rate. Before reaching the cellular concentration at about 6x105cells/ml the cell line was in lag phase. The lag time was measured as the time taken for the cells to exceed their initial plating density and the population doubling time was estimated at the midpoint of the exponential phase of the growth curve. The values were found to be 1 and 5 days respectively (Fig. 2).
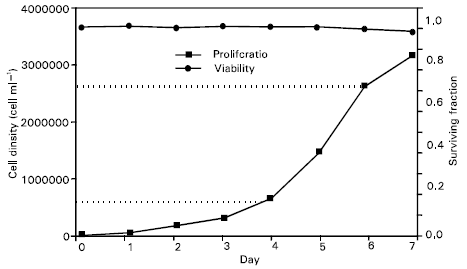 | |
| Fig. 1: | Jurkat T cell line: proliferation and viability during 7-day long culture in an environment of 37oC temperature, 80% humidity and 5% CO2. The seeding or initial cell density was taken 3.7x104 cells/ml, and no further dilution was done during the experiment. The log phase interval for the cells is indicated by the two horizontal dotted lines |
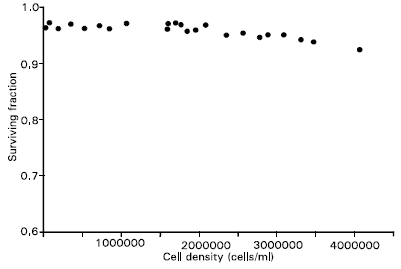 | |
| Fig. 2: | Dependence of the fraction of surviving cells of Jurkat on cell density in culture |
The fraction of surviving cells of Jurkat on cell density in culture show that the viability of cells remained within the range 96-97% up to cell density about 2.7x106 cells ml–1. At very higher cell density, the viability was slightly decreasing. This may be due to the unavailability of medium or nutrients for the cells because as cell density increases cells become more tightly packed together and as a result, the surface area available for nutrient uptake becomes smaller. A given concentration of nutrients can only support a certain number of cell (Griffiths, 1979). A cell can often find alternative nutrients when one becomes exhausted, but this is not actually beneficial for the cell viability because the growth rate is always reduced. Nutrients become growth limiting before they become exhausted. As the concentration of amino acids falls, the cell finds it increasingly difficult to maintain sufficient intracellular pool levels.
The dose-response curves for inactivation of Jurkat cells incubated for 4 hours with 1.0 mM ALA at different temperatures (10-50°C) were studied. A typical example of the dose-responses at temperatures 10-37°C has been shown Fig. 3. The light exposure following 4 hours incubation with 1.0 mM ALA induced a phototoxic effect for the cells. The phototoxic effect first increased very slowly and then increased rapidly with increasing light dose. The light exposure for 50% cell survival, D50, was estimated to be 7.2 min, which corresponded to the total irradiance value of 1.04 W/cm2 because the energy flux at cell position was 2.5 mW/cm2. The cell survival for incubation temperature of 31°C did not follow the exponentially decaying, as expected in PDT. This survival is modeled as a sigmoidal Boltzman function (curve) with constant parameters shown in figure as follows:
| (1) |
It can be seen that the dose-responses and cell viability incubated at 10, 15, 20, 25, 33, 35 and 37°C show almost the same fashion (not shown).
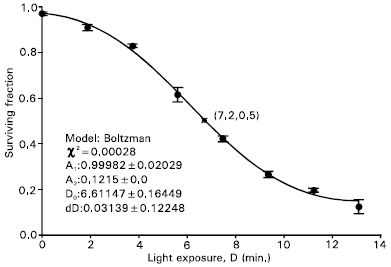 | |
| Fig. 3: | Dose-response curve for inactivation of Jurkat cells incubated for 4 hours at 31°C with 1.0 mM ALA in 80% humidity and 5% CO2. Samples of cells proliferating in the exponential phase with cell densities 1-2x106 cells/ml were chosen. Bars, SD from six independent experiments. When bars are not shown the SD are smaller than the symbols. A sigmoidal (Boltzman) curve (with constant parameters shown in figure): ySF = A2 + (A1-A2)/(1 + exp((D-D0)/dD)) is fitted to the data. The light exposure for 50% survival, D50, is shown by a star mark and is 7.2 min, which corresponds to the irradiance value of 1.04 W/cm2 |
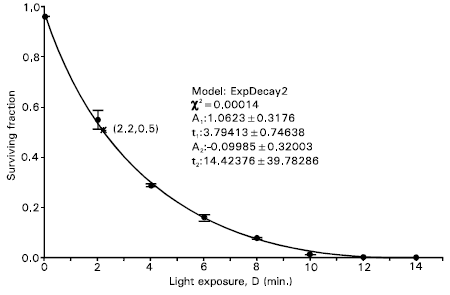 | |
| Fig. 4: | Dose-response curve for inactivation of Jurkat cells incubated for 4 hours at 43°C with 1.0 mM ALA in 80% humidity and 5% CO2. Bars, SD from three experiments. A second-order exponential decay curve (with decay constants and other constant parameters shown in figure): ySF = A1 exp(-D/t1) + A2 exp(-D/t2) is fitted to the data. The D50 is 2.2 min |
These are not still exponentially decaying, and are modeled as the same sigmoidal Boltzman curve as for 31°C (equation 1) with varying constant parameters. For each temperature a shoulder at the lower exposure region of the sigmoidal curve was obtained. The phototoxic effect within shoulder is very small, which means that a minimum exposure time (or light dose) is required to introduce the cell inactivation process or overcoming the threshold reaction; low temperatures have delayed the process. The length of shoulder was found to decrease with the increase in temperature, which indicated that at higher temperatures the cellular damage had been occurred promptly.
The cell survivals for 39, 41, 43, 45 and 50°C temperatures were found to follow the second-ordered exponential decay with varying decay constants and constant parameters. The model for these concave or tailed surviving functions is:
| (2) |
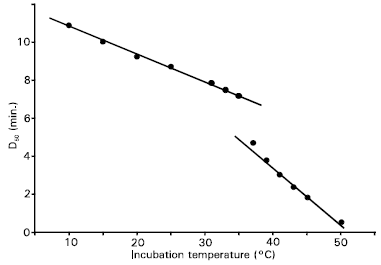 | |
| Fig. 5: | Dependence of light exposure for 50% cell survival, D50, on incubation temperature. Two linear curves with different slopes are fitted to the data |
With increasing incubation temperature the tail of the exponential survival curve was found to lengthen. As an example, a dose-response curve for inactivation of the cells incubated at 43°C temperature has been shown Fig. 4. The viability of cells with zero light dose remained within the range 96-97% in the incubation temperature range from 10-50°C, with a maximum at 35-40°C. The D50 for each incubation temperature was calculated. The dependence of the D50 on incubation temperature is shown in Fig. 5. As seen in the figure, two linear curves with different slopes are fitted to the data. The fittings show that at lower incubation temperatures for which the cell viability follows the sigmoidal Boltzman curve has lower slope than that at higher temperatures for which the viability follows the exponential decay. However, the overall dependence of the light exposure for 50% survival on temperature is linearly decreasing. This means that the efficiency of cell inactivation is higher at higher temperatures, in general. The fitting with higher slope confined the data points within the temperature range from 39°C to 50°C. The higher slope obtained at higher temperatures indicates that at this temperature range the rate or efficiency of cell inactivation is rapid. This may be due to the contributions from heating effects for cell constituents and nutrients or cell surrounding's medium as well as from the accumulation of PPIX, as a photosensitizer, from ALA in the cell system (Moan et al., 1999; Moan et al., 1998; Gottfried et al., 1991 and Patrice et al., 1979). While ALA itself is not a photosensitizer, it can induce the biochemical formation of PPIX via the biosynthesis process, which is actually part of the overall scheme for the production of a biologically useful form of chemical energy. Every cell in the body requires energy for metabolism, and since most nucleated cells obtain energy via a process that involves heme-containing enzymes, they must have at least a minimal capacity to synthesize PPIX. Under normal conditions, heme biosynthesis is regulated so closely that photosensitizing concentrations of PPIX never accumulate except in a few highly specialized tissues. A substantial portion of the PPIX biosynthetic process is centered in the mitochondria and is under strict enzymatic control. Since the cellular concentration of ALA is regulated by an enzymatic feedback control involving ALA synthase, it is possible to produce large but transient amount of PPIX by bypassing the ALA synthase rate-limiting step. Though the penetration of ALA into the cells is practically not temperature dependent, the production of PPIX is strongly temperature dependent (Moan et al., 1999; Moan et al., 1998 and Juzenas et al., 1999). ALA-induced PPIX production in cells have been reported by Dietel et al. (1996). They declared that when cell temperature was raised to 35-40°C, the rate of porphyrin formation was increased for both PPIX and water-soluble uroporphyrin and coproporphyrin at incubation period of 3 hours, and for 6 hours incubation periods, the production of the water-soluble porphyrins was continued in a quasi-linear manner with respect to the incubation time. Gottfried and Kimel (1991) have studied temperature effects on photosensitized reactions and have shown that in the temperature range 15-45°C a twofold increase or more in reaction rate has been observed on chemical acceptors. Patrice et al. (1979) demonstrated that the temperature during hematoporphyrin derivative treatment greatly influenced the photocytotoxicity on both normal progenitors and leukemia cells. They reported that temperature variation induced changes in cell metabolism. Because PPIX is a photosensitizer, the efficacy of ALA for use in PDT depends on the accumulation of sufficient levels of PPIX in cells (Juzenas et al., 1999 and Melø et al., 1985). Low temperatures are likely to result in a higher pH or less cell metabolism and thus decreased drug uptake. Both of these effects will decrease cell killing. Cellular sensitivity to PDT-induced toxicity is known to be influenced by a number of factors. These include the oxygenation of the cells (Mitchell et al., 1985), the tolerance of the cell to free radicals, which is enhanced if excess ferric ion is present (Lin et al., 1995), and the intercellular concentration of PPIX (Iinuma et al., 1994), which depends on the balance between the rate of PPIX synthesis and its removal by cellular eflux and conversion to heme.
In conclusion, proliferation studies for the cells showed that for the cell densities larger than about 2.7x106 cells/ml, after 6 days of culture, the proliferation of the cells was remained constant, indicating the culture in the plateau phase. For the cell density smaller than 6x105 cells/ml the culture was in lag phase. This indicates that the culture within the cellular concentration from 6x105-2.7x106 cells/ml was in exponential or log phase, where the viability and reproduction for the cells were higher, which might be used for sampling the cell population. The lag time and proliferation time were found to be 1 and 5 days respectively. From the cell density dependent viability, it was found that the viability for cell density up to 2.7x106 cells/ml did not depend on the density of the cells in culture, whereas at very higher cell densities, it was found to decrease slightly. The viability of cells remained within the range 96-97% in the temperature range about 10-50°C. The cell was found to be more viable at temperatures 35-40°C. The dose-response curves for incubation temperatures 10-37°C were modeled as sigmoidal Boltzmann curves, whereas the dose-response curves for temperatures 39-50°C were modeled as the second-order exponential decay curves. The efficiency for cell inactivation was found to increase with increasing incubation temperature. The higher production rate of photosensitizing PPIX at higher temperatures might play an important role in the observed higher cytotoxic effect at higher incubation temperatures.
ACKNOWLEDGMENT
The Norwegian Government under the Quota program supported this work.