
D.A. M. Choudhury
Entomology Division, Bangladesh Rice Research Institute, Gazipur-1701, Bangladesh
M.J. W. Copland
Department of Agricultural Sciences, Imperial College at Wye, Wye, Ashford, Kent TN25 5AH, UK
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 5 | Page No.: 455-460







ABSTRACT
Trichome types and density on the abaxial surface of five host plants of glasshouse leafhopper Hauptidia maroccana Melichar were studied by scanning electron microscopeic (SEM) procedure. The movement of the egg parasitoid Anagrus atomus Linnaeus was also studied on five different host plants. Glandular and non-glandular erect trichomes were observed on the leaf surfaces. Trichome density was always higher on leaf veins than on leaf lamina. The highest (348 mm-2) and lowest (6 mm-2) trichome density was observed on Primula and Sweet pepper, respectively. Variation in leaf morphology of different host plants significantly affected the walking speed and turning of A. atomus. Walking speed of female A. atomus was the fastest (1.21 mm sec-1) on Sweet pepper and slowest (0.72 mm sec-1) on Primula which attributed to the highest meandering (55.48 deg mm-1) on Primula and the lowest (21.61 deg mm-1) on Sweet pepper among the five host plants. On the other hand, the presence of host egg worked as arrestant. The females showed frequent turning and walked most slowly in presence of host egg in the leaf. Consideration should be made on plant surface structures in determining the release of A. atomus in biological control programme.
PDF Abstract XML References Citation
How to cite this article
D.A. M. Choudhury and M.J. W. Copland, 2003. Influence of Plant Structural Complexity on the Searching Behaviour of the Egg Parasitoid Anagrus atomus (Linnaeus) (Hymenoptera: Mymaridae). Pakistan Journal of Biological Sciences, 6: 455-460.
DOI: 10.3923/pjbs.2003.455.460
URL: https://scialert.net/abstract/?doi=pjbs.2003.455.460
DOI: 10.3923/pjbs.2003.455.460
URL: https://scialert.net/abstract/?doi=pjbs.2003.455.460
INTRODUCTION
Chemical and physical characteristics of the plant can mediate the behaviour of parasitoids and predators of herbivores (Eigenbrode and Espelie, 1995). Plant structural complexity like cuticular thickness, glandular and non-glandular trichomes can play a significant role in host encounter by parasitoid wasps (Dicke, 1996; Lukianchuk and Smith, 1997). When a parasitoid lands on a plant, plant surface characteristics, mainly trichomes alter their walking speed (Hulspas-Jordaan and Van Lenteren, 1978), inhibit their ability to locate a host (Treacy et al., 1985; Keller, 1987; Kashyap et al., 1991) and finally reduce adult survival (Obrycki and Tauber, 1984). Plant volatiles can acts as repellents and trichome and trichome exudates inhibit the movement of the wasp on the plant (Romeis et al., 1998).
Plants with trichomes and their secretions can impede the movement of parasitoids, often affecting searching time and weakening the parasitoid response (Sütterlin and Van Lenteren, 1997; Romeis et al., 1999; Clercq et al., 2000; Lovinger et al., 2000). Efficiency of several parasitoid species were affected negatively a high trichome densities. A higher walking speed of Encarsia formosa was observed on a hairless cucumber variety than on a variety with intermediate trichome density (Van Lenteren et al., 1995). The searching behaviour of egg parasitoid Trichogramma exiguum Pinto and Platner was found to be affected by surface chemistry among different plant species (Keller, 1987). Thus understanding the natural enemy response to host plant resistance characteristics is important for successful integration of tactics used in pest management.
Hauptidia maroccana is a polyphagus insect and feed and lay eggs on wide ranges of host plants with a variety of trichomes and semiochemicals. Anagrus are considered polyphagus parasitoids, yet many are limited to a few plant species although some host leafhoppers have a broad plant range. For example, Stilling (1980) found high rates of parasitism for an Anagrus species associated with three Eupteryx leafhopper species on stinging nettle, but a little or no parasitism on surrounding weedy vegetation containing Eupterys. Chantarasa-ard et al. (1984) found that Anagrus incarnatus Haliday parasitized Harmalia albicollis Motschulsky eggs on rice in the laboratory, but they did not detect parasitism in the field when host eggs were deposited in weeds. These examples indicate that Anagrus species may have an inherent attraction to specific plant volatiles. Since the searching efficiencies of predators and parasitoids is influenced by the presence of trichomes and the semichemicals in the host plants, the success of biological control may be influenced by the characteristics of different host plants. The present study was undertaken to identify the characteristic difference of the host plant surface on the basis of types and density of the trichomes and to determine the effect of physical characteristics on searching efficiency of Anagrus atomus on five different host plants.
MATERIALS AND METHODS
Scanning electron microscope (SEM) studies of the abaxial surface of five host plant species: A scanning electron microscope study of the following five host plants of H. maroccana was done to find the characteristic differences in the abaxial surfaces of the leaf:
| • | Cucumber, Cucumis sativus L. (var. Bush champion) |
| • | Tomato, Lycopersicon esculentum Mill. (var. Alisa Graig) |
| • | French bean, Phaseolus vulgaris L. (var. Prince) |
| • | Primula, Primula vulgaris Huds. (var. Mixed colour) and |
| • | Sweet pepper, Capsicum annum L. (var. Worldbeater) |
Fully expanded leaves of the above mentioned host plants were collected from the glasshouse. The samples were put inside a plastic box and immediately transferred to the electron microscopy unit for the preparation of specimens. The preparation of specimens consisted of the following stages:
Drying: In the electron microscope, specimens were exposed to a very high vacuum. This could use the water-containing specimens to dry out and collapse. To prevent this, biological specimens are usually dried in a special way, which minimized distortion.
Fixation: After drying, the specimen was immersed in a chemical fixative solution, which killed the cells and preserved them. Glutaraldehyde, mixed with a phosphate buffer, is a common fixative. Samples of 10 mm across, were cut carefully from the leaves without damaging them. They were immersed in the 2.5% Glutaraldehyde in phosphate buffers for twelve hours. After this period the specimens were placed in two changes of phosphate buffer alone for ten minutes to remove surplus fixatives.
Dehydration: During this process, water was gradually removed from the specimen and replaced with acetone. The specimen was dipped in 50% acetone for 20 min, in 70% acetone for 20 min or left in the fridge overnight, in 80% acetone for 20 min, in 90% acetone for twenty min, in 100% acetone for 20 min, in 100% acetone for 20 min, in 100% acetone for 20 min.
Critical point drying: The specimen was sealed in a 'critical point drying apparatus'. Here acetone was replaced with liquid CO2. The temperature and pressure were then increased until the 'critical point' for CO2 is reached. Under this condition CO2 was a vapour. In fact, while the temperature and pressure were rising, the specimen was gradually drying, but in such a way that no damaging surface tension forces occured within it.
The critical point dryer specimen container is filled with 100% acetone. The specimens were transferred to the container quickly to avoid desiccation. The drying process tooks a minimum of two hours. On completion the specimens must be kept dry and dust free.
Attachment to stub: An adhesive such as Araldite® or a special colloidal silver mixture was used to mount the specimen securely on a metal stub. That acted as a specimen holder in the SEM.
Gold-coating: The specimen, adhesive and stub were coated with a very thin layer of gold in a ‘sputter coater’. That was necessary because otherwise an electrical charge would build up within the specimen while it was being examined in the SEM. Instead, gold conducts the charge away to earth.
The gold-coated specimens were loaded into a (Hitachi SS430) scanning electron microscope and observed at different magnifications. The photographs were taken by a built in camera at various magnifications.
Searching behaviour of A. atomus on the abaxial surface of the leaves of five host plant species: The behavioure differences during searching for hosts by A. atomus on five different host plants were observed under laboratory condition at room temperature (25±1°C). Fifty mm diameter petri-dish cages were used in the experiment. Leaf discs of 50 mm diameter from the healthy plants were placed on 1% agar medium facing the lower surface uppermost. Twenty four hours old mated, unexperienced female were used in this experiment. The wasps were provided with 50% honey solution prior to using in the experiment. Single female wasp was released onto the leaf disc.
Different activities of the parasitoid was monitored through a JVC camera with a 50 mm macro lens, attached to the computer and parasitoid behaviour was recorded in video tapes using the computer programme ‘Inchworm’ (Varley et al., 1994). Parasitoid behaviour on each leaf disc was recorded for 30 min. Moreover, the behaviour of A. atomus was also recorded on P. vulgaris leaf disc containing H. maroccana eggs to find whether the presence of host altered its searching behaviour. Recording was carried out between 0900-1400 h to avoid any diurnal variations. Cold light was provided with fibre optic at 55.5 W m–2, with both arms placed 10-12 cm over the arena. The same parasitoid was used in different leaves and considered as one replication but each leaf disc was used only once. Ten replications were done for each plant species.
The videotapes were replayed and the direction of walking of the parasitoid was traced by clicking on the right mouse button. Any parasitoid stayed at least 15 seconds continuously on the leaf surface was accepted as one replication. The computer programme 'Inchworm' gave the following information:
| • | Net speed (mm sec–1) |
| • | Meandering (degrees turned mm–1) |
| • | Absolute turn (degrees turned sec–1) |
| • | Statistical analysis |
One-way analysis of variance was done in SAS to compare different activities during searching by A. atomus in different host plant leaves and the leaves of P. vulgaris containing eggs of H. maroccana. Activities on leaf lamina and leaf veins on different host plants were compared using two sample t-tests in MINITAB. Means were compared in LSD at 5% level.
RESULTS AND DISCUSSION
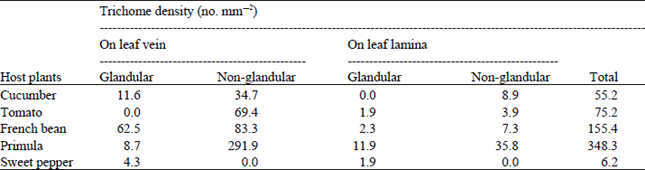
Structural differences of the abaxial surfaces of host plant leaves: A great variation has been observed in the types, density and the arrangements of trichomes on the abaxial surface of the leaves of five plant species on the basis of the SEM study. Trichome density was always higher on leaf veins than on leaf lamina. Among the five plant species, the highest (348.3 mm–2) and lowest (6.2 mm–2) trichome density was observed in Primula and Sweet pepper, respectively (Table 1).
Both glandular and non-glandular erect trichomes were found on the leaf lamina of Cucumber leaf but glandular trichomes were absent on leaf veins. Density of non-glandular trichomes in Cucumber was observed as 3.9 and 69.4 trichomes per mm2 on leaf lamina and leaf veins, respectively. Shipp and Whitfield (1991) observed 489.3 trichomes on a 4 cm diameter leaf disc of Cucumis sativus L. ‘Sandra’.
Both erect and glandular trichomes were found in Primula, 85% of which were non-glandular. The head of the glandular trichomes were single celled. Trichomes were densely arranged on both leaf lamina and leaf veins.
The abaxial surface of French bean leaves was uniformly covered with glandular and hooked trichomes at almost equal proportions on both leaf lamina and leaf veins.
Both glandular and non-glandular trichomes were observed in Tomato leaves. The head of the glandular trichomes was four celled. In Tomato, 97% of the trichomes were non-glandular. Leite et al. (1999) also mentioned 90.3 and 9.7% of non-glandular and glandular trichomes, respectively in Tomato.
Only glandular trichomes were found in Sweet pepper and the trichomes were present only on leaf lamina. The head of the trichomes was two celled. Shipp and Whitfield (1991) observed 16.9 trichomes on a 4 cm diameter leaf disc of Capsicum annum L. ‘Plutona’.
The dense trichomes on the abaxial surface of leaves can provide protection against herbivore attack in many ways. The hooked trichomes in French bean can catch the arthropods, immobilizing them and causing death from dehydration and starvation before they can escape. On the other hand, the glandular trchomes, which may protects the plant from herbivorous attacks secrete sticky exudates when ruptured.
The glandular trichomes found in Tomato have four-celled head. These trichomes are known as type A Gibson’s is common in various Solanaceous species and they release sticky exudates when ruptured by insects (Jeffree, 1986). These highly specialized secretory cells synthesize and accumulate a large variety of terpene oils and other essential oils whereas the principal toxic component has been identified as 2-tridecanone (Dimock and Kennedy, 1983).
| Table 1: | Trichome density on vein and on leaf lamina of the five different host plants under SEM study |
 | |
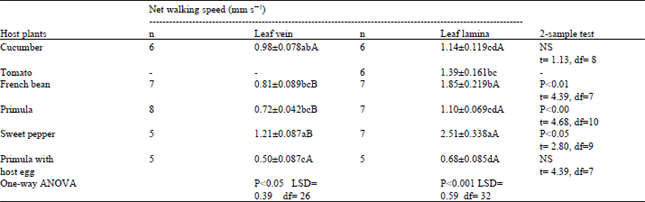
| Table 2: | Net walking speed of the egg parasitoid Anagrus atomus on leaf surface of different host plants |
 | |
| Table 3: | Absolute turning (degrees sec–1) of the egg parasitoid Anagrus atomus during searching for eggs on the surface of different host plants |
 | |
| Table 4: | Meandering (degrees mm–1) of the egg parasitoid A. atomus during searching for eggs on the surface of different host plants |
 | |
| Means in columns, followed by the same lowercase and in rows followed by the same uppercase letters are not significantly different from each other at P≤ 0.05. One-way ANOVA followed by Least Significant Difference test, n = No. of insects as replicates | |
Searching behaviour of A. atomus on the undersurface of the leaves of five host plant species: When the parasitoids were introduced into the leaf disc without host eggs they started to move in any directions, changed direction frequently and apparently randomly, sometimes reaching the petri-dish wall. They then walked along the side wall and then again would change direction. But when leaves contain host eggs parasitoids started drumming with their antennae immediately after introducing into the leaf discs and never left the leaves. Plant features such as leaf toughness or hairiness, or increased trichome density on leaves, make the parasitoid less effective in finding hosts (Van Driesche and Bellows, 1996). One of the characteristics that can negatively influence the behaviour, is the presence of leaf hairs (Kleijn et al., 1997). The results of the present study indicate that leaf surfaces of different host plants influenced walking speeds and turning of the egg parasitoid A. atomus. The variation in the walking behaviour on the leaf surface of different plant species may be due to the morphological differences in leaf structure, particularly trichome forms and density.
Significant difference was observed in the net walking speed of the parasitoid on the leaf vein (P<0.05) and leaf lamina (P<0.001) of five plant species. Net speed was the highest (1.21 and 2.51 mm s–1) on Sweet pepper and lowest (0.72 and 1.1 mm s–1) on Primula along the leaf vein and leaf lamina, respectively (Table 2). Net speed was lower in leaf containing host eggs than on clean leaf. Between the two areas of the leaf, the net speed was always higher on leaf lamina than on leaf veins in five host plant species.
No significant difference was observed in the absolute turning (degrees s–1) of the parasitoid on leaf vein and leaf lamina of five different host plants and even when the leaves contained host eggs (Table 3). The 2-sample t test did not show any significant difference in absolute turning between leaf lamina and leaf veins on five host plant species.
Meandering (degrees mm–1) of the parasitoid showed a significant difference on different plant species both on leaf vein (P<0.001) and leaf lamina (P<0.001) among the five host plants. The highest and lowest meandering was observed on Primula and Sweet pepper, respectively. Moreover, the turning was higher on the leaf containing host eggs than on the clean leaf (Table 4).
The differences in the walking speed might be due to the variation in trichome density on different plant species, which prevented the parasitoid to walk through a straight path and led to more turning. These results agree with the findings of Sütterlin and Van Lenteren (1997) who observed the highest walking speed (0.73 mm s–1) of Encarsia Formosa Gahan on hairless Sweet pepper leaves and the lowest on Cucumber (0.2 mm s–1) with stiff and large hairs in a high density. Van Lenteren et al. (1995) had also mentioned a higher walking speed of E. formosa on hairless (0.60 mm s–1) than on hairy (0.40 mm s–1) cultivar of Cucumber. This may illustrate that leaf hairiness in general hampers parasitoid activity.
The presence of glandular trichomes of leaf surface can adversely affect the searching behaviour of parasitoid wasps. Lovinger et al. (2000) reported that the egg parasitoid A. nigriventris Girault, spent less time and showed fewer oviposition probes, tended to fly off the stems of alfalfa with glandular trichomes than on alfalfa without glandular trichomes. Romeis et al. (1998) mentioned that the movement of Trichogramma chilonis Ishii pigeon pea pods and calyxes was inhibited by long trichomes and were trapped by sticky trichome exudates. In another study Romeis et al. (1999) observed T. chilonis to spend a longer time on chickpea leaves when the acidic trichome exudate was washed off than on unwashed leaves.
Anagrus atomus stayed most of the time either on the lid or on the side of the petri-dish when they were introduced in to Tomato leaf discs. The parasitoid wasp never walked over the leaf veins and walked fast on the leaf lamina and fled to the lid of the petri-dish. This might be due to the presence of secondary plant metabolite ‘Tomatin’ or the glandular trichome exudates that repelled the parasitoid wasp. Farrar and Kennedy (1991) found that the glandular trichome exudates of the resistant wild Tomato Lycopersicon hirsutum glabratum, which contains 2-tridecanone acted as a repellent and harmed the egg parasitoid Telenomus sphingis (Ashmead) at high concentrations, could entangle it, repeled it or reduced its searching efficiency or even killed the parasitoid by poisoning.
Parasitoid behaviour was reported to change with the presence of hosts on a leaf. The egg parasitoid A. nigriventris Girault spent more time on plants containing host eggs and the proportion of fast walking time was significantly greater on plants without host eggs (Al-Wahaibi and Walker, 2000). The parasitoid Encarsia formosa stayed longer on leaves where they could recognize hosts or host products (Van Lenteren et al., 1996). In the present study, A. atomus walked slowly and made more turns per mm on the leaves containing host eggs.
The results of the present study indicated that structural variation of plant surfaces significantly affected the searching rates of A. atomus on different plant species. This means that habitat structure influences the searching success of a parasitoid and it should be considered during application of a biological control programme. However, the results of laboratory studies may not directly predict the magnitude of effects of plant surface characteristics on parasitoid behaviour in the field. More laboratory as well as field studies are needed to confirm the predicted effects of leaf surfaces and parasitoid age on oviposition behaviour and the rate of parasitism by A. atomus.
REFERENCES
- Shipp, J.L. and G.H. Whitfield, 1991. Functional response of the predatory mite Amblyseius cucumeris (Acar: Phytoseiidae), on western flower thirps, Frankliniella occidentalis (Thysanoptera: Thripidae). Environ. Entomol., 2: 649-699.
Direct Link - Van Lenteren, J.C., Z.H. Li, J.W. Kamerman and R. Xu, 1995. The parasite-host relationship between Encarsia formosa (Hym: Aphilinidae) and Trialeurodes vaporariorum (Hom: Aleurididae) XXVI. Leaf hairs reduce the capacity of Encarsia to control greenhouse whitefly in cucumber. J. Applied Entomol., 119: 553-559.