
Haque N. M. Mozaddedul
Entomology Division, Bangladesh Rice Research Institute, Gazipur -1 701, Bangladesh
M.J. W. Copland
Entomology Section, Department of Agricultural Sciences, Imperial College at Wye,
University of London, Wye, Ashford, Kent TN25 SAH, UK
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 4 | Page No.: 422-431







ABSTRACT
Searching behaviour of the parasitoid, Leptomastix nr. epona were evaluated on four different leaf surfaces (Passiflora, Avocado, Guava, Citrus) and the arrestment behaviour of the parasitoid to the honeydew and waxes of Pseudococcus longispinus and P. affinis were compared under a video camera in laboratory. Scanning electron microscopy were also done on four leaf surfaces. No significant differences were observed in the behaviour of the parasitoid, L. nr. epona towards the honeydew of long-tailed (host) and glasshouse mealybug (non-host). Female Leptomastix nr. epona immediately after release, starts searching in clean, honeydew and waxes contaminated areas. After contacting with honeydew or wax contaminated areas, the parasitoid searches the area intensively with its antenna and then turn around the honeydew or waxes contaminated area and then try to probe by its ovipositor. The female parasitoids may consider long-tailed mealybug honeydew and waxes as chemical and visual cues and responds to this stimulant by changes in locomotary behaviour. Overall, the present results showed L. nr. epona searches on honeydew or wax coated areas more than on a clean area. So honeydew and wax might act as an arrestment stimulus or visual cue, showing this associated host product is a long-term kairomone. The net speed of searching of Leptomastix nr. epona on Passiflora was reduced compared with other three species and the number of trichomes observed on Passiflora (leaf lamina and mid rib of the leaf) were higher compared with other three plant species. So it may be one of the reason of the parasitoid low net speed of searching on dense trichomatous leaf lamina such as Passiflora compared to leaf lamina where no trichome at all (citrus).
PDF Abstract XML References Citation
How to cite this article
Haque N. M. Mozaddedul and M.J. W. Copland, 2003. Searching and Arrestment Behaviour of the Parasitoid,
Leptomastix nr. epona (Walker) as Affected by Leaf Surfaces, Honeydew
and Waxes of Mealybugs. Pakistan Journal of Biological Sciences, 6: 422-431.
DOI: 10.3923/pjbs.2003.422.431
URL: https://scialert.net/abstract/?doi=pjbs.2003.422.431
DOI: 10.3923/pjbs.2003.422.431
URL: https://scialert.net/abstract/?doi=pjbs.2003.422.431
INTRODUCTION
Searching behaviour is an active movement by which insects seek resources. It is an important kind of behaviour because insects thereby acquire food, mates, oviposition sites and refugia (Bell, 1990). When the female parasitoid reaches the proper host; this step is recognized as ‘host location’ (Vinson, 1975 and 1976; Vinson and Iwantsch, 1980; Van Alphen and Vet, 1986). A parasitoid may cue to its host’s food plant utilising factors different from those used by the host in locating a plant. Thus a host may have a wide range of plants on which it develops, but the parasitoid may only respond to some of these plants but not others (Vinson, 1981).
Plants may produce chemicals that attract and retain parasitoids or provide nutrition (Shahjahan, 1974) and thereby attract parasitoids (Williams et al., 1988). It has been known for long while that plant play a role in host location by parasitoids and an increasing number of studies have identified specific plant volatiles that attract the parasitic wasps to them (Quicke, 1997). Any attempt to understand the factors involved in population changes of an insect pest would be incomplete if the relationship between plant and insect is excluded.
Several plant characters are known to affect the searching behaviour and parasitization efficiency of Trichogramma spp. (Hymenoptera:Trichogrammatidae) (Romeis et al., 1999). Keller (1987) mentioned that plant surface characteristics might affect walking parasitoids by altering walking speeds. Physical aspects of plants affect the natural enemies in several ways. Spatial dispersion of plants can affect the ability of herbivores and natural enemies to locate and in some cases successfully colonize the plants. Plant features such as leaf toughness or hairiness, which in some cases defend plants against herbivores, may also affect natural enemies. Increase trichome density on leaves influences the walking speed and lowers the success rates of foraging hosts (Van Driesche and Bellows, 1996).
Chemicals appear to play a major role at almost every level of the host selection process. Plant volatiles emanating from the host’s food or food plant and food odours have been shown to be important cues in host habitat location for a number of Hymenopterous as well as several of the dipterous parasitoids (Williams et al., 1988). The parasitoid will often land and examine the damaged plant tissue with her antennae (Doutt, 1959). Such behavioural changes may be caused by plant-liberated factors operating at a short range. If a non-host causes the tissue damage, the parasitoid may resume searching (Vinson and Lewis, 1965). However, if the damage is due to the host, the parasitoid’s behaviour is changed and she excitedly walks over the plant or contaminated surface, rubbing the substrate, with her antennae and appearing to search for the host (Williams, 1951; Vinson and Lewis, 1965; Vinson, 1968; Lewis, 1970). Romeis et al. (1999) cited that leaf chemicals could mediate the behaviour of parasitoids and predators of herbivores. Plant compounds are used directly by natural enemies as cues for habitat location in some instances. In other instances, plant compounds may either be sequestered by herbivores or may be released by plants under herbivore attack, resulting in the attraction of natural enemies directly to the host (Van Driesche and Bellows, 1996).
Parasitoids often home in on volatile chemicals associated with a host frass, symbiotic fungi, or damage to plants caused by host feeding (Quicke, 1997). There is a considerable variety in the source of these chemicals, which are called ‘kairomones’ (Waage, 1979). A kairomone is defined as a transpecific chemical messenger, the adaptive benefit of which falls on the recipient rather than on the emitter. These chemicals include the vast number of attractants, phagostimulants and other substances that mediate the positive responses of predators to their prey, herbivores to their food plants and parasites to their hosts (Brown et al., 1970). Honeydew is a reliable indicator of the presence of a host and has been shown to be an important source of kairomones for many aphid and coccid parasitoids (Quicke, 1997). Bouchard and Cloutier (1984) suggested that some odours given off by aphids or by honeydew could act as a general attractant. The same authors found honeydew as a source of host searching kairomones for aphid parasitoid Aphidius nigripes Ashmead. Carter and Dixon (1984) found that the response of coccinellid larvae to honeydew is similar to the response of parasitoids to the contact chemicals of their hosts. These authors reported honeydew as an arrestant stimulus for coccinellids. Budenberg (1990) also reported honeydew as a contact kairomone for aphid parasitoids. Once parasitoids have found a microhabitat, searching for hosts is often guided by responses to chemical stimuli deposited by the host or caused by the host’s presence. Parasitoids use many chemical cues during host selection; the known sources of chemical stimuli include host frass, mandibular gland secretion, host cuticle and accessory gland secretion (Ambriz et al., 1996). The host plant characteristics and its innate behavioural pattern govern parasitoid searching time, pattern and efficiency. The searching efficiency is hampered by length, density and alignment (erect, procumbent, etc) of the leaf pubescence. Insects may be trapped by sticky exudate of glandular trichomes, leading to slower walking speed and poor encounter rates. Vertically oriented long trichome impair normal searching patterns and sometimes irritate the insect leading to it abandoning the surface and flying away from the prey. Tingle and Copland (1988) noticed that the degree of parasitism varies with the plant species. Therefore, the studies were carried out-to determine whether the long-tailed mealybug honeydew mediates host location in the parasitioid, Leptomastix nr. epona as a kairomone, to find out if there are any differences in preferences for host (P. longispinus) and non-host (P. affinis) honeydew, to investigate the arrangement of trichomes of a number of host plants using scanning electron microscopy (SEM), to investigate the effects of leaf trichomes and plant volatiles on parasitoid host searching behaviour.
MATERIALS AND METHODS
Arrestment behaviour of the parasitoid, Leptomastix nr. epona to the honeydew of long-tailed and glasshouse mealybug-: The arrestment behaviour of the parasitoid was observed under a video camera. The experimental patches consist of a circular disc of Passiflora coccinea leaf (50-60 mm diameter) placed in Petri-dishes containing a thin layer of agar gel (8 g l-1) to maintain high humidity. Freshly collected honeydew of P. longispinus and P. affinis were smeared on each half portion of the leaf disc of Passiflora coccinea individually. The patch was then offered to an experienced, mated and 4-8 days old female parasitoid, which were fed with 50% honey solution for a period of 24 hours prior to use in the experiment. Each female wasp was released into a covered arena and its activities were recorded on videotape. The magnification was adjusted until the arena filled the screen. Light was provided by a fibre optic cold light source at 45 W/m2, with both arms placed 10-12cm over the arena. Room temperature was 23-24oC. The arrestment behaviour of the parasitoid, Leptomastix nr. epona was observed for 15 minutes. Twelve parasitoids were observed.
Arrestment behaviour of the parasitoid, Leptomastix nr. epona to an area coated with honeydew and waxes of the long-tailed mealybug and to a clean area: The procedure was the same as the last experiment except twelve’s mealybugs (size class-3 and size class-4) were placed on half of the area for 24 h whilst the other half was kept clean (by covering with aluminium foil). After 24 h the mealybugs were removed from the arena and the experiment was carried out. Fifteen replications were observed.
Scanning electron microscopy (SEM) of different leaf lamina of four plant species: Citrus (Citrus limon) (Rutaceae), Passiflora (Passiflora coccinea) (Passifloraceae) Avocado (Persea americana) and Guava (Psidium guajava) (Myrtaceae) were selected for that purposes. Uniformity of the experimental leaves was maintained by taking a fully mature 5th/6th leaf from the terminal bud. The leaves were uninfected and samples were collected just before the experiment and were used without rubbing or soaking in water, which may alter natural surface characters of the leaves. Only complete leaves were taken to avoid the possible effect of damage surfaces. They were removed from the plants with the help of scissors and forceps, placed in small plastic box on a wet blue tissue and immediately transferred to the microscopic unit for the preparation of specimens. The specimens (leaf samples) were prepared in three stages I. e. drying, attachment to stub and gold coating. The gold-coated specimens were loaded into a Hitachi S430 Scanning Electron Microscope and observed at different magnifications. A built-in camera at various magnifications took the photographs.
Searching behaviour of the parasitoid, Leptomastix nr. epona on different leaf lamina of four plant species-: Uniform size, 6-8 days old female parasitoids were collected from the culture. Experimental patches consisted of a freshly excised circular leaf disc (50-60 mm diameter) of each of four plant species (i.e. I. Citrus limon, ii. Passiflora coccinea, iii. Persea americana, iv. Psidium guajava) placed in a Petri-dish containing a thin layer of agar gel (8 g/litre) to maintain humidity. The patch of each plant species was offered to an experienced, mated and 6-8 days old female parasitoid (which was fed with 50% honey solution for 24 h prior to experiment) one after another individually for behavioural observation. Each female wasp was released into a covered arena and its activities were recorded on videotape. The magnification was adjusted (before) until the arena filled the video screen. Light was provided by fibre optic cold light source at 45 W/m2, with both arms placed 10-12 cm over the arena. Room temperature was 24-25oC. A test run was carried out to obtain a suitable magnification and appropriate colour to distinguish leaf surface from the test insect in arena. After standardization, the position of the camera was fixed throughout the experimental period. The parasitoid was individually introduced into the arena with the leaf to be searched for 15 minutes. Each female parasitoid and each leaf surface was used only once. Ten fresh leaves per plant species served as replicates. The movements of the parasitoid was monitored through a JVC camera with a 50 mm macro lens, attached to a computer which analyzed the movements using the computer program ‘Inchworm’ (Varley et al., 1994) and gave the following information with regard to searching:
Resting (stopping) time: This is the time the parasitoid stops during searching. It is measured in seconds.
Net speed: The net speed is the actual speed calculated after excluding the stopping time from the total time. It is measured in millimetres per second.
Degrees turning (meandering) per millimetre: The mean turns/distance (degrees turned/mm moved)
Degrees (Absolute turns) turning per second: The mean turns/time (degrees turned/second)
Data recording procedure: The introduced insect was allowed to settle down onto the leaf surface and then the stopwatch and the video recorder were started simultaneously. Any insects, which stayed more than 15 seconds on the experimental leaf lamina, were considered as replicates. Recording was carried out between 0900-1400 h to avoid any diurnal variation. The data were analysed in two way ANOVA by using GENSTAT 5.1 and comparisons were made using LSD at 5% levels.
RESULTS
Arrestment behaviour of the parasitoid, Leptomastix nr. epona to the honeydew of long-tailed and glasshouse mealybug-: The arrestment behaviour of the parasitoid to the honeydew of P. longispinus and P. affinis were compared. The parasitoids spent most of their time on the walls of the arena (Fig. 1). There were no significant differences observed in the behaviour of the parasitoid towards the honeydew of either mealybugs (Fig. 2).
Arrestment behaviour of the parasitoid, Leptomastix nr. epona to an area coated with honeydew and waxes of the long-tailed mealybug and to a clean area: Once the parasitoids, were released into the arena, they promptly started searching on the Passiflora leaf.
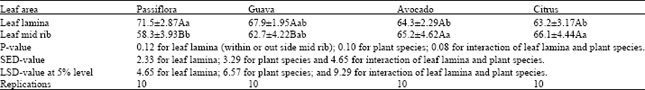
| Table 1: | Searching behaviour of the parasitoid, Leptomastix nr.epona on a clean area and area coated with honeydew and waxes of P. longispinus |
 | |
| 1 Time spent in seconds by searching ± SE. 2 Net distance travelled/second (millimetres) ± SE. | |
| Table 2: | Trichome number per mm2 on different leaf position of four plant species under SEM |
 | |
| Table 3: | Searching behaviour (Degrees turning per second ± s.e.) of the parasitoid, Leptomastix nr.epona on the leaf lamina and mid rib of four plant species |
 | |
| Table 4: | Searching behaviour (Degrees turning per millimetre ± s.e.) of the parasitoid, Leptomastix nr.epona on the leaf lamina and mid rib of four plant species |
 | |
| Table 5: | Searching behaviour (Net speed ± SE.) of the parasitoid, Leptomastix nr.epona on leaf lamina and mid rib of four plant species |
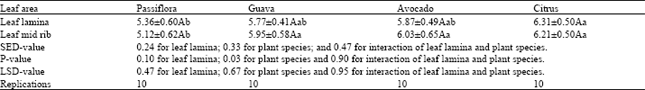 | |
| Net speed = Net distance travelled/second (millimetres). Means in columns followed by same uppercase letters are not significantly different at P= 0.05 (LSD). Means in rows followed by same lowercase letters are not significantly different at P= 0.05 (LSD). Two ways ANOVA were carried out. | |
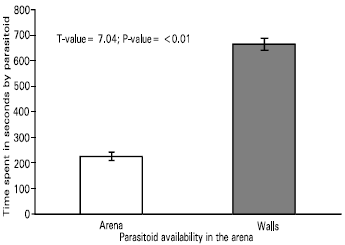 | |
| Fig. 1: | Time spent in the arena or on the walls during a searching bout of 900s by the parasitoid, Leptomastix nr. epona to the honeydew of Pseudococcus longispinus and Pseudococcus affinis, n = 15 |
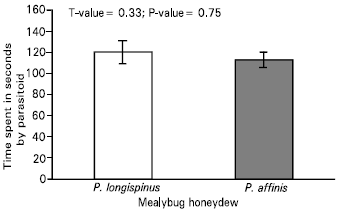 | |
| Fig. 2: | Time allocation of the parasitoid, Leptomastix nr. epona to the honeydew of Pseudococcus longispinus and Pseudococcus affinis in a choice test, n = 15 |
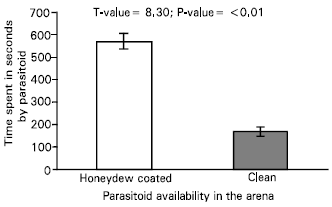 | |
| Fig. 3: | Time spent by the parasitoid, Leptomastix nr. epona in the area coated with honeydew and waxes of Pseudococcus logispinus and clean area during a searching bouts of 900s. n = 15 |
 | |
| Fig. 4: | Time spent by the parasitoid, Leptomastix nr. epona in the area coated with honeydew and waxes of Pseudococcus longispinus and clean area during a searching bout of 900s. n = 15 |
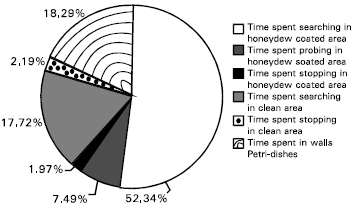 | |
| Fig. 5: | Percent time spent of the parasitoid, Leptomastix nr. epona to the long-tailed mealybugs honeydew and wax coated area and clean area. n = 15 |
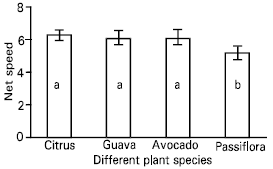 | |
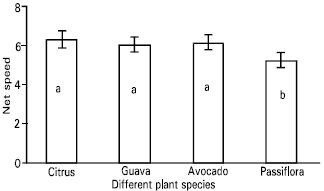
| Fig. 6: | Net speed [ distance travelled (millimetres)/second] of the parasitoid, Leptomastix nr.epona on leaf lamina of four plant species (Citrus, Guava, Avocado and Passiflora). n = 10; bar = SE of mean |
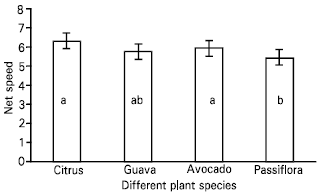 | |
| Fig. 7: | Net speed [distance travelled (millimetres)/second] of the parasitoid, Leptomastix nr.epona on mid rib of four plant species (Citrus, Guava, Avocado and Passiflora). n = 10; bar = SE of mean |
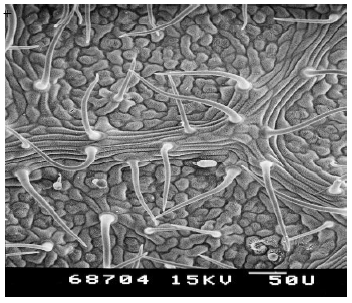 | |
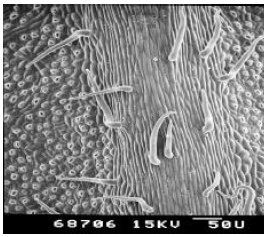
| Plate 1: | SEM of the under-side (leaf lamina) of passiflora (Passiflora coccinea) leaf |
 | |
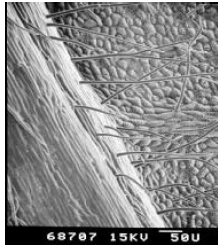
| Plate 2: | SEM of the under-side (mid rib of the leaf) of passiflora (Passiflora coccinea) leaf |
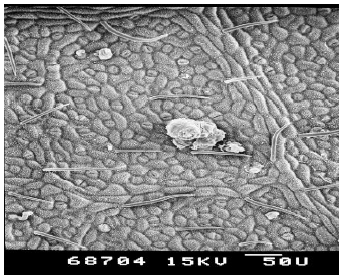 | |
| Plate 3: | SEM of the under-side (leaf lamina) of avocado (Persea americana) leaf |
 | |
| Plate 4: | SEM of the under-side (mid rib of the leaf) of avocado (Persea americana) leaf |
 | |
| Plate 5: | SEM of the under-side (leaf lamina) of citrus (Citrus limon) leaf |
 | |
| Plate 6: | SEM of the under-side (mid rib of the leaf) of citrus (Citrus limon) leaf |
The parasitoids spent more time searching in the arena than on the walls (Fig. 4). The time spent by the parasitoids in the area on the leaf coated with honeydew and waxes were significantly different from clean area (Fig. 3). On encountering fragments of wax the parasitoids turned around and antennated the site several times and probed (mean number 3.10±s.e. 0.91) with their ovipositor. After probing the parasitoid examines the probed area again by its antennal contact and tries again to probe.
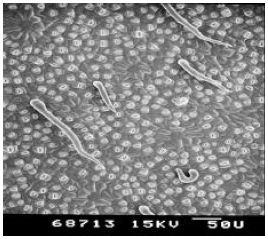 | |
| Plate 7: | SEM of the under-side (leaf lamina) of guava (Psidium guava) leaf |
 | |
| Plate 8: | SEM of the under-side (mid rib of the leaf) of guava (Psidium guava) leaf |
The parasitoid performs an arrestment response on the substrate; she walks slowly and the frequency of turning increases. The female regularly repeats these behavioural sequences after a stop and preens herself. Once a female wasp loses contact with the contaminated zone, she usually returns quickly towards the area. The presence of honeydew causes several different mechanisms including arrestment in the parasitoid, a change in walking speed (orthokinesis) and an increase in the rate of turning (klinokinesis) (Table 1). Time spent by the parasitoid in different activities is presented in Fig. 5. Among different activities of the parasitoid, percent time spent in searching on honeydew coated areas was higher (52.34%) compared with clean areas (17.72%). Analysis of different searching parameters revealed that parasitoids respond to an area coated with honeydew and waxes of the mealybugs by a slower walking speed, a longer time spent in searching, greater degrees of turn for each unit distance walked and degrees of turn per seconds compared with clean area.
Time spent in searching was significantly different on the two areas (Table 1).
SEM of different leaf lamina of four plant species: On the basis of the SEMs obtained, the structure of the under surfaces of four-plant leaf lamina were categorised as: (a) glossy, (b) glabrous and (c) pubescent. The individual characteristics of each plant leaf lamina and mid rib of the leaf are discussed as follows:
Passiflora coccinea: A perennial climber with grooved downy stems, leaves 70-150 cm long and 30-70 cm wide rounded to subcorate at base, irregularly toothed, 2-6 glanded.
The under-side of the leaf lamina was densely pubescent. Many shapes of trichomes were noted including erect, pointed, hooked, slightly hooked and glandular. Trichome number was higher on mid rib of the leaf compared to leaf lamina. Stomata were very small and not protruding from the leaf lamina (Plates 1 & 2).
Persea americana: The under-side of the leaf lamina was glabrous but rough. Trichomes were pointed, slightly hooked and sparse, located along side veins. Thichome number was higher on midrib of the leaf than leaf lamina. Stomata were small and abundant and uniformly distributed all over the leaf lamina (Plates 3 & 4).
Citrus limon: The under-side of the leaf lamina was glossy and smooth. No trichomes were present on either the leaf lamina or mid rib of the leaf. Stomata were small and distributed abundantly across leaf lamina but very few on mid rib of the leaf (Plates 5 & 6).
Psidium guajava: The under-side of the leaf lamina was densely pubescent. Many shapes of trichomes were noted including erect, pointed, hooked and slightly hooked. Trichome number was higher on mid rib of the leaf than leaf lamina. Stomata were small and protruding from the leaf lamina (Plates 7 & 8).
The trichome number per mm2 of the four plant species were higher on the midrib compared with the leaf lamina except on Citrus (where no trichomes were present) (Table 2).
Searching behaviour of the parasitoid, Leptomastix nr. epona on different leaf lamina of four plant species-: After parasitoids were released onto the leaf lamina, they immediately started exploring the leaf lamina or midrib of the leaf. The wasps were not constrained and could leave the leaf lamina at any time. The parasitoid moved in any direction, changed direction frequently and apparently randomly until reaching the Petri-dish wall. They then walked alongside the wall and then again would change the direction.
Significant differences were observed in the degrees turning per second of the parasitoid, L. nr. epona on leaf lamina and midrib of Passiflora and Guava compared with other plant species and between the leaf lamina and midrib of Passiflora and Guava (Table 3).
Significant differences were also observed in the degrees turning per millimetre of the parasitoid on leaf lamina of Passiflora compared with the other plant species and between the leaf lamina and mid rib of Passiflora and Avocado (Table 4).
A significant slower net speed of the parasitoid was observed on Passiflora compared with other three species (Table 5, Fig. 6 and Fig. 7).
DISCUSSION
The role of honeydew and waxes in the attraction of the parasitoid, L. nr. epona: Host-derived components of larval feeding damage and frass will be important cues to which innate responses may exist (Vet et al., 1995). Parasitoids locate their host through a variety of physical and chemical cues, the most important of which are factors associated directly with host such as feeding secretions and frass (Vet et al., 1991). During the process of host experience the parasitoid learns several things associated with the host (Tumlinson et al., 1992), including the odour of chemical cues that are produced by it and/or because of its presence in that particular environment.
The results showed that there were no significant differences observed in the behaviour of the parasitoid, L. nr. epona towards the honeydew of long-tailed (host) and glasshouse mealybug (non-host). But female of Leptomastix nr. epona immediately after release, started searching in clean, honeydew and wax contaminated areas. After contact with honeydew or wax contaminated areas, the parasitoid searches the area intensively with its antenna and then turns around within the honeydew or wax contaminated areas and tries to probe with her ovipositor. This study suggests that female parasitoids may consider long-tailed mealybug honeydew and waxes as chemical and visual cues and responds to this stimulant by changes in locomotary behaviour. Further investigations are needed to determine if L. nr epona is able to distinguish between the volatiles released by infested plants either with or without the presence of host and host by-products and whether volatiles are systematically released by damaged plants.
Similarly Mehrnejad (1998) reported that both experienced and inexperienced parasitoids of Psyllaephagus pistaciae responded well and were arrested in the honeydew (A. pistaciae) contaminated patches. Vet et al. (1995) suggested nalive parasitoids are expected to have a potential to respond innately to the stimuli derived from their host, from the food and environment of the host and from organisms associated with the host.
During the studies, the parasitoid, Leptomastix nr. epona on encountering fragments of wax or honeydew turns around and antennates the site several times and probed with its ovipositor. This finding clearly indicates that volatile compounds associated with honeydew act as a cue in the host finding process of L. nr. epona. Similarly Shaltiel and Ayal (1998) reported that the wasp Diaeretiella rapae uses honeydew emitted by its host, the cabbage aphid Brevicoryne brassicae, as a kairomone. Shimron et al. (1992) concluded that arrestment by honeydew is usually indicative of host presence, moreover this increases the efficiency of host location, placing the parasitoid in a better position for reproductive success.
The parasitoid, Leptomastix nr. epona spent more time in honeydew or wax-coated areas than clean areas. This may be due to the presence of the kairomone in the host honeydew and is certainly advantageous for the parasitoid since such areas are the most likely to contain long-tailed mealybugs. Many homopterous species excrete honeydew on which other insects feed. In their review Hagvar and Hofsvang (1991) reported that many studies have shown increased searching times in honeydew contaminated areas by aphid parasitoids. This indicates that honeydew is a host location kairomone for Aphididae as well as a source of food. The parasitoid, Leptomastix nr. epona utilizes long-tailed mealybugs honeydew as a source of food. It could be questioned that the positive responses found here might be an attempt to feed, but this is unlikely because the parasitoid used in the experiments had been fed with 50% honey solution prior to experiments. Similarly, Mehrnejad (1998) reported that the parasitoid, P. pistaciae utilises psyllid honeydew as a source of food and he also found positive responses to honeydew in his experiment.
The response of L. nr. epona to honeydew from P. affinis (non-host) and P. longispinus (host) suggests that intensive search of areas in which host mealybugs were not present could occur. However, the response to honeydew is probably additive to the response to the food plant of their mealybug hosts and therefore, although some time could be spent searching on inappropriate plants it is likely to be a small proportion of the overall time spent searching by the parasitoid. The long-tailed mealybug, P. longispinus colonies produce a relative small amount of honeydew that accumulates on infested leaves, twigs, fruits, flowers and branches and the surrounding surface and remains for a short period. But waxes of P. longispinus remain for a long time. It is expected that honeydew increases the parasitoids host searching efficiency in the areas in which mealybugs occur, moreover host honeydew is a very good food source for parasitoids.
Effect of leaf surface properties on searching behaviour of the parasitoid: The plant physical structure and its surface affect host-plant selection. The hairs and wax-layer on the surface of the leaf influence the movements and feeding of insects Physical aspects of the plants affect the natural enemies in several ways. Spatial dispersion of plants can affect the ability of herbivores and natural enemies to locate and in some cases successfully colonize the plants (Southwood, 1986). Plant features such as leaf toughness or hairiness, which in some cases defend plants against herbivores, may also affect natural enemies. Increases in trichome density on leaves is associated with slower walking speed and lower rates of foraging, making the parasitoids less effective at finding hosts (Van Driesche and Bellows, 1996).
The net searching speed of Leptomastix nr. epona on Passiflora (Leaf lamina and mid rib of the leaf) was reduced compared with other three plant species (avocado, guava and citrus). This may be due to the number of trichomes on Passiflora (leaf lamina and midrib of the leaf) which were high in comparison other three plant species. Similarly Heidari (1989) reported that good control of Pseudococcus viburni (Signoret) was achieved by Cryptolaemus montrouzieri (Mulsant) irrespective of the hairness of the plant species, but Nephus reunioni Fürsch was more effective on smooth leaves, such as those of citrus, coffee and Passiflora, than on hairy plants, such as Streptocarpus and tomato, on which the density of the trichomes and their sticky exudations significantly reduced the walking speed and other searching parameters.
Gillani (1998) reported that the plant leaf surfaces had a significant effect on the net searching speed of the lacewing, Symphorbius fallax and he observed maximum net speed on citrus and minimum on guava and he also observed that net speed on smooth leaves such as citrus, coffee, mango and yam was high as compared to that on hairy leaves, i.e. guava.
The net speed of searching of the parasitoid, Leptomastix nr. epona low on Passiflora leaf lamina and mid rib compared with three other plant species leaf lamina and mid ribs (citrus, avocado and guava) and also observed on scanning electron micrographs (SEMs) that the number of trichomes were higher on passiflora, compared to citrus, avocado and guava (Plates 7-8). So this may be one of the reasons for the parasitoids low net speed of searching on dense trichomatous leaf lamina such as Passiflora compared to leaf lamina where there are no trichomes at all (citrus). Since P. longispinus is found on a variety of host plants, the leaf surface topography, i.e. presence or absence of trichomes and their intensity, leaf arrangement, can affect the searching efficiency of L. nr. epona.
ACKNOWLEDGMENTS
The authors thank to Sue Stickels and Jon Varley for their technical help and the Bangladesh Rice Research Institute for funding the research. We would like to thank Bas Nijhof in the Netherlands who provided the parasitoid. We also thank Chris Hodgson at Imperial College at Wye, University of London and U.K. for identification of the mealybug species. Our thanks are extended to Alan Clewer for statistical advice on the design and data analysis of the experiments. All work was carried out at Imperial College at Wye.
REFERENCES
- Bell, W.J., 1990. Searching behavior patterns in insects. Ann. Rev. Entomol., 35: 447-476.
CrossRefDirect Link - Hagvar, E.B. and T. Hofsvang, 1991. Aphid parasitoids (Hymenoptera, Aphidiidae): biology, host selection and use in biological control. Biocontrol News Inform., 12: 13-42.
Direct Link - Vinson, S.B. and G.F. Iwantsch, 1980. Host suitability for insect parasitoids. Annu. Rev. Entomol., 25: 397-419.
CrossRef