
Ilham M. El- Rafai
Botany Department, Faculty of Science, Tanta University, Tanta, Egypt
Susan. M.W. Asswah
Botany Department, Faculty of Science, Tanta University, Tanta, Egypt
Omaima. A. Awdalla
Botany Department, Faculty of Science, Tanta University, Tanta, Egypt
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 4 | Page No.: 399-406







ABSTRACT
Four biocontrol–agents, namely : Trichoderma harzianum, T. hamatum, Bacillus subtilis and Pseudomonas fluorescens, have been tested for their potential antagonism for controlling fusarium wilt, verticillium wilt and early blight diseases of tomato. In vitro studies showed that culture filtrates of all antagonistic organisms significantly decrease the spore germination and germ tube-length of the tested pathogens, F. oxysporum f. sp. lycopersici, Verticillium dahliae and Alternaria solani. The linear growth and sporulation of the concerned pathogens were also inhibited the degree of inhibition was varied according to the tested antagonistic filtrate. In vivo studies, three treatments were applied; inoculation of the soil with antagonist period to sowing, soaking tomato seeds in the filtrate of the tested antagonist before sowing and coating of tomato seeds with spores of the antagonist before planting. Soil inoculation and seed coating with T. hamatum spores completely controlled the concerned diseases and improved the yield. However, P. fluorescens seed coating controlled the early blight disease and improved the tomato growth as well. Concerning the chemical assessment, T. hamatum soil inoculation and seed coating treatments gave the highest increase for chlorophyll a, b and cartenoids. Also the same treatments showed the highest increase of phenolic compounds (free and conjugated) and the lowest percentage for sugars content of tomato leaves infected with the concerned pathogens.
PDF Abstract XML References Citation
How to cite this article
Ilham M. El- Rafai, Susan. M.W. Asswah and Omaima. A. Awdalla, 2003. Biocontrol of Some Tomato Disease Using Some Antagonistic Microorganisms. Pakistan Journal of Biological Sciences, 6: 399-406.
DOI: 10.3923/pjbs.2003.399.406
URL: https://scialert.net/abstract/?doi=pjbs.2003.399.406
DOI: 10.3923/pjbs.2003.399.406
URL: https://scialert.net/abstract/?doi=pjbs.2003.399.406
Introduction
The widespread use of fungicides to control plant diseases has led to an increase of health hazards due to their phytotoxic residual and pollution effects, therefore using some other means of disease control instead of agrochemical is strongly encourage.
The biological control of plant pathogenic fungi has received considerable attention as an alternative strategy. The use of the antagonistic properties of Trichoderma spp in the biological control of many plant diseases has been a subject of many studies (Dubos et al., 1978; Papavizas, 1985; Bicici et al., 1991; Wolffhechel and Jensen, 1992; Lumsden, 1993; Awad, 1995; Ushamalini et al., 1997; El- Shanshoury et al., 2000 and Osman et al., 2001). In all these studies, antibiosis was potentially a principal component of mechanism of the biocontrol by Trichoderma spp, which produced an array of metabolites identified as antifungal and antibacterial compounds (Abd El Moity, 1981; Chisalberti and Sivasithamparan, 1991).
Antagonistic bacteria have been extensively studied as biocontrol agents effective against various soil – borne pathogens. Among 20 genera of bacteria, Bacillus spp, Pseudomonas spp and streptomyces spp are widely used as biocontrol agents. Several Bacillus spp including B. subtilis are antagobistic to plant pathogenic fungi and bacteria. Bacillus spp produced at least 66 different antibiotic compounds (Ferreira et al., 1991). Subtilin, bacillin, bacillomycen, subtenolin, mycosubtilin, toximycin and bacitracin are different names given to antibiotics produced and Bacillus subtilis (Schober 1984; and Loeffler et al., 1986). The antagonistic effect Bacillus subtilis against several fungi In vitro and in vivo have bean reported by (Utkhede and Rahe, 1980; Wolk and Sarkar, 1994; Saleh, 1997; and El-kazzaz et al., 2000). However, investigation on mechanisms of biological control by Pseudomonas spp, revealed that P. fluorescens produced an antibiotics highly effective which protect plants from various pathogens through inducing systemic resistance (Gurusiddaiah et al., 1986; Frommel et al., 1991 and Zhou and Paulitz, 1994).
Tomato (Lycoperiscon esculentum Mill) is susceptible to many diseases that reduces its yield. The most serious of which are the wilts and early blight caused by Fusarium oxysporum f. sp. lycopersici, Verticillium dahliae and Alternaria solani, respectively. The present investigation aimed at studying the effects of four antagonistic microorganisms on growth activities of some tomato pathogenic fungi in vitro. The second objective was to evaluate the efficiency of different application treatments of these microorganism in reducing disease incidence., and their effect on plant growth. Finally to assess biochemical changes associated with disease development after treatment with these antagonistic microorganism.
MATERIALS AND METHODS
Microorganisms and culture conditions: Three isolates of fungi pathogenic to tomato, namely, Fusarium oxysporum f. sp. lycopersici (sacc.) snyder a Hansen, causing fusarium wilt, Verticillum dahliae (C. strak.) causing verticillium wilt and Altrenaria solani (Ell. and Mart.) Jones and Grout, causing early blight were used in the present study. Fungal isolates were maintained as single hyphal tips on Czapek- Dox agar. Isolation of bioagents used in the present study were carried out using rhizosphere soil samples collected from tomato fields of El–Gharbia Governorte , Egypt.
Preliminary screening revealed that Trichoderma harzianum, Trichoderma hamatum, Bacillus subtilis and Pseudomonas fluorescens were the most antagonistic microorganisms against the above mentioned pathogenic fungi. Trichoderma spp were grown on (gliotoxin fermentation medium) for 7 days at 27°C according to (Brain and Hemming, 1945). Bacillus subtilin was grown for 3 days on (liquid nutrient glucose broth) at 30°C under complete darkness to stimulate toxin production (Dowson, 1957). On the other hand Pseudomonas fluorescens was grown for 3 days at 25°C on (King’s medium B) according to (King et al.,1959).
Host plant and soil: Seeds to tomato cultivar (Lycoperiscon esculentum Mill C.V Moneymaker) were used throughout this study. Seeds were surface- sterilized by soaking in 5% sodium phypochlorite solution for 2 minutes and then washed thoroughly in a sterile water. The soil used in the green house experiment was a sandy clay loam. Soil was air-dried, sieved through a 2-mm sieve and mixed fine sand at a ratio 2: 1 (w/w).
Effect of culture filtrates of the tested antagonistic on growth activities of pathogenic fungi in vitro: Each of the aforementioned bioagent isolates was grown in its previously mentioned medium, then 10 ml sterile distilled water was added to the substrate mycelium. The mixture was homogenized and filtered through whatman No. I paper and the filtrate was sterilized through 0.4 μm filter. The method described by El-Abyad et al. (1983) was used. A standard agar disc (5 mm) of each of the tested tomato pathogen grown on modified Czapek-Dox medium for 10 days was transferred aseptically to a vial containing 10 ml of each antagonistic sterile filtrate. The vials were shaken vigorously for 5 min to suspend the macroconidia. Vials containing sterile distilled water were served as control. One ml spore suspension of each fungus in each antagonistic filtrate and control was pipetted onto microscope slide in petri dishes. The plates were incubated at 27°C for 10 hours for F. oxysporum f. sp. lycopersici and V. dahliae. On the other hand A. solani was incubated for 15 hours as determined in preliminary experiment. The percentage of germination was calculated in 100 spores from 5 fields selected at random. The length of germ tubes was also estimated recorded and the average was calculated. Three slides were prepared for each treatment and the means were recorded.
Linear growth and sporulation: Aliquots of Czapek-Dox media were prepared containing the different anatgoistic filtrate and control containing no filtrate. After the poured plates were set each was inoculated with a 5 mm diameter disc cut from a young active culture of each tested tomato pathogen. Three replicates (plates) for each antagonistic filtrate were incubated in the dark for a period of 6 days at 27°C, after which the colony diameters were measured. Sporulation was studied by inoculating Czapek-Dox agar plates amended with different sterilized antagonistic filtrates with a 5mm disk of each tested fungus. The plates were incubated for 10 days at 27°C. At the end of incubation period, 1 cm2 agar discs were cut from the margin of the colony and transferred to a vial containing 10 ml of sterile distilled water, the suspension was continuously shaken for 5 min, after which time, the density of spores/ml was counted by a haemocytometer according to El- Abyad et al. (1983). Three plates were used for each treatment and the mean number of spores was calculated.
Preparation of antagonistic inocula: Each suspension of each antagonistic was prepared as previously mentioned and used for the following treatment 1-soil pre-inoculation, in which antagonistic suspension was adjusted to contain 3 x 106 C.F.U ml and mixed with talc powder at the rate of 1:1 (v/w), then dried for 3 days at room temperature to produce dry powder according to (Abd El Moity, 1981) 2- Seed soaking treatment , in which tomato seeds were soaked separately in the filtrate of each tested antagonistic (3x106 C.F.U ml) or distilled water as control. Drench was done for 2 hours period to sowing 3-Seed coating, in this treatment, tomato seeds were coated with colloidal suspension containing (3 g of each antagonistic strain + 1 g of sodium alginate) or distilled water as control. The treated seeds were left of dry about 3 hours before planting.
Soil infestation: Each pathogenic fungus was grown on Czapek’s Dox both in Erlenmeyer flasks and incubated at 27°C for 7 days. The mycelia of each fungus was harvested on whatman No. 1 filter paper, then resuspended in sterile distilled water and blended for 30 sec, after which the appropriate volume for the soil treatment was obtained. The pathogenic fungi were inoculated into the soil at a rate of 2% of the soil weight (Rothrock and Gottlieb, 1984).
Biocontrol treatments (in vivo): The experiments were carried out in plastic pots (20 cm in diameter containing 1 kg of soil adjusted to 60% of its moisture holding capacity. Twenty seeds were sown in each pot and three pots were used for each treatment in each infested soil. In the soil pre- inoculation treatment powder of each tested antagonistic were added to soil just before planting at the rate 5 g pot-1. Meanwhile treated seeds (soaked and coated) were sown at the same time of soil pathogen inoculation,. The pots were kept at 27± 3°C, watered daily and fertilized weekly. The infection percentage and dry weight of plants were recorded 3, 6 and 9 weeks after sowing respectively. In all treatments both of pathogen free soil and untreated infested soil served as control.
Chemical assessment: Photosynthetic pigments i.e chlorophyll a and b as well as carotenoids were determined in treated plants and controls photometrically according to the method described by Nornal (1982). All determination were calculated as mg g-1 fresh plant materials. Phenolic and sugar contents were also determined as follows, ethanolic extracts of leaves (healthy and diseased) were used for determination of both phenolics (free, conjugated and total phenols)and sugar contents (reducing, non– reducing and total sugars) according to Snell and Snell, (1953) and Forsee, (1938).
Statistical analysis: The result obtained were subjected to statistical according Snedecor and Cochran, (1989).
RESULTS AND DISCUSSION
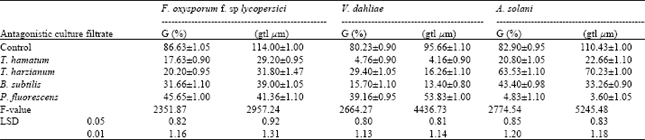
Effect of culture filtrates of antagonistic microorganisms on growth activities of the tested tomato pathogens (in vitro): Data of microscopic studies presented in (Table 1, 2) indicate that both of the spore germination percentage and germ tube-length of the tested pathogens (Fusarium oxysporum f. sp. lycopersici, Verticiltiun dahliae and Alternaria solani) were significantly decreased by treatment with culture filtrate of the tested antagonistic. The inhibitory effect varied according to the antagonistic. The filtrate of T. hamatum was more effective on the three tested pathogen, while B. subtilis was very effective on V. dahliea and P. fluorescens had a deleterious effect on A. solani more than other experimental antagonistic filtrate. (Table 1).
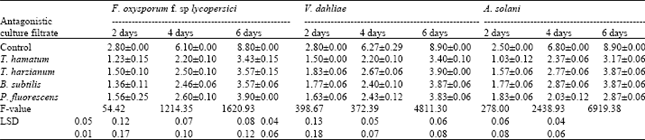
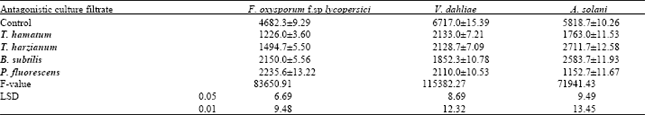
Results in (Table 2) indicates that the culture filtrates of antagonistic significantly inhibited the radial growth of the tested pathogens. The degree of inhibition was varied according to the antagonistic filtrates. T. hamatum was the most effective one in inhibiting the radial growth comparable on the others. The data obtained in (Table 3) show that the number of spores of the tested pathogenic fungi was significantly decreased by the antagonistic filtrates of all tested isolates. Sporulation of the tested pathogenic fungi varied according to the source of the antagonistic filtrate.
The microscopic study revealed that the culture filtrate of the antagonistic organisms, Trichoderma harzianum, T. hamatum, Bacillus subtilis and Pseudomonas fluorescens significantly decreased the conidial germination, germ tube-length, radial growth and sporulation of the tested pathogens. The suppressive effect varied according to the antagonistic filtrate. The present investigation indicate that T. hamatum was the more effective among the others. These results were in accordance with (Chet and Baker, 1981; Bicici et al., 1991 and Ushamalini et al., 1997) Trichoderma spp is known to have the ability to produce some extracellular lytic enzymes that are involved in the process of antagonism against a variety of pathogenic organisms (Mukhapadhyay, 1997). The positive effect of T. hamatum against the tested tomato pathogens could be explained by hyperpararsitism. The results obtained in the present work indicate that B. subtilis has clear antagonistic effect against the tested pathogens. The present results are in harmony with the work of Abd El Moniem, (1996) who found that an isolate of B. subtilis has antagonistic effect against S. rolfsii In vitro and in vivo. In the meantime Saleh (1997) found that the B. subtilis significantly decreased the incidence of root rot and wilt of ground nut. Asaka and Shoda (1998) suggested that the antagonistic activity of B. subtilis against several host fungi in vitro may be referred to the production of the antibiotics such as iturin A and surfactin. In the present study P. fluorescens proved to have antagonistic activity against the tested pathogens. The suppressive effect of P. fluorescens is due to the production of phenazine –1- carboxilic acid (a product of the shikimic acid pathway). This antibiotics is effective against several fungi (Gurusiddaiah et al., 1986).
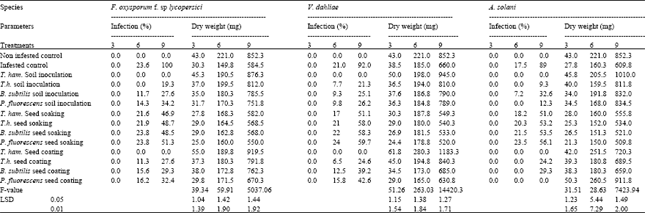
Biocontrol treatment (in vivo): Data presented in Table 4 indicated that all the tested biocontrol agents were significantly differed in its effect on percentage infection and yield. Moreover, there were significant differences among the tested tomato pathogens. Table 4 shows that both of soil pre-inoculation and seed coat treatment with T. hamatum completely prevented the infection of tomato with F. oxysporum f. sp. lycopersici.
| Table 1: | Effects of culture filtrate of T. hamatum, T. harzianum, B. subtilis and P. fluorescens on spore germination (G%) and average germ tube length (gtl Fm) of F. oxysporum f. sp lycopersici, V. dahliae and A. solani |
 | |
| Table 2: | Effects of culture filtrate of T. hamatum, T. harzianum, B. subtilis and P. fluorescens on linear growth (mm) of F. oxysporum f. sp lycopersici, V. dahliae and A. solani after 10 days incubation at 27 °C |
 | |
| Table 3: | Effects of culture filtrate of T. hamatum, T. harzianum, B. subtilis and P. fluorescens on sporulation of F. oxysporum f. sp lycopersici, V. dahliae and A. solani after 10 days incubation at 27°C |
 | |
| All F-values are significant at 0.0001 | |
Infestation of soil with F. oxysporum f. sp. lycopersici alone, decreased growth yield. However, seed coating with T. hamatum was the best treatment for the control of fusarium wilt and improving the plant growth. Table 4 indicates that soil pre-inoculation and seed coating with T. hamatum was very effective in controlling verticillium wilt caused by V. dahliae as compared with the seed soaking treatment. Dry weight was significantly increased in seed coating treatment. Results obtained in Table 4 show that no infection symptoms of early blight of tomato caused by A. solani was observed for 9 weeks after the soil pre inoculation with T. hamatum as well as P. fluorescens seed coating treatments. Dry weight was significantly increased in T. hamatum soil pre inoculation treatment as compared with other treatments and control. In vitro studies evaluated the beneficial effect of these experimental microorganisms on the wilt and early blight tomato pathogens. These results stimulated the evaluation of the antagonistic activity In vivo. This was achieved by using different treatments. The first was inoculation of the soil with the antagonist in the form of powder prior to sowing. The second, was drench in which the seeds were soaked in the filtrates for certain time before sowing. The third, was coating tomato seeds with the spores of antagonist before planting. The obtained results indicate that control of pathogens and plant growth of tomato differed according to the treatment and the antagonistic organisms. The most effective treatment for controlling all tested pathogens was seed-coating. Similar results have been obtained by other workers in the control of several diseases, maize root rot (Singh and Mehrotra, 1980); Fusarium root rot of faba bean (Yehia et al., 1982); damping off of sugar beet (Martin and Hancock, 1987) fungal and bacterial diseases of tomato (El-Abyad et al., 1993). In all these studies and in the present study, plant growth was improved by seed – coating treatment.
| Table 4: | Effects of different treatments with the tested antagonist on tomato fusarium wilt, verticillium wilt, and early blight at different cultivation periods |
 | |
| Table 5: | Effects of different treatments with the tested antagonist on photosynthetic pigments, chlorophyll a, chlorophyll b, and carotenoid (mg g-1 fresh weight of leaves) of tomato infected with F. oxysporum f. sp lycopersici, V. dahliae and A. solani at different cultivation periods |
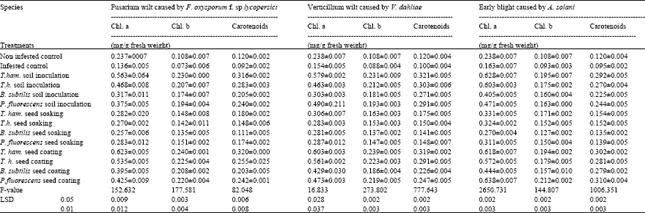 | |
| All F-values are significant at 0.0001, T. ham= Trichoderma hamatum, T.h= Trichoderma harzianum, B. subtilis= Bacillus subtilis, P. fluorescens= Pseudomonas fluorescens | |
| Table 6: | Effects of different treatments with the tested antagonist on phenol and sugar of tomato infested with F. oxysporum f. sp lycopersici, V. dahliae and A. solani |
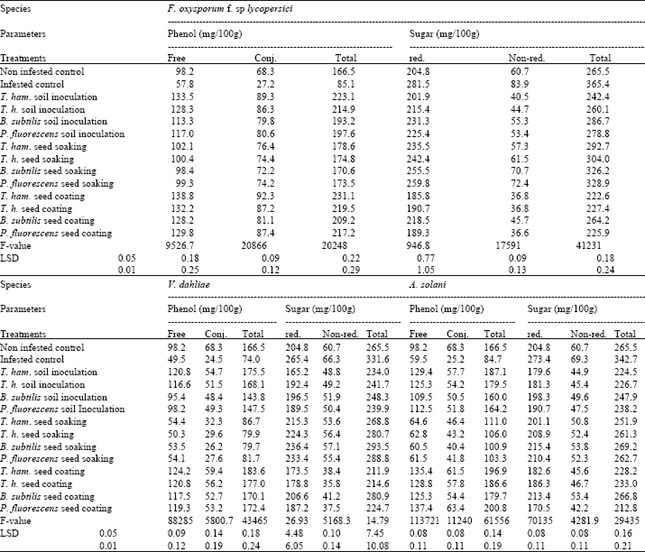 | |
| All F-values are significant at 0.0001, T. ham= Trichoderma hamatum, T. h= Trichoderma harzianum, B. subtilis= Bacillus subtilis, P. fluorescens= Pseudomonas fluorescens | |
The present study showed that seed- soaking treatment was the least effective treatment in controlling the tested pathogens. This may be attributed to tomato hard seed coat, which decreased the absorption capacity of tomato seeds to the antagonist filtrates (El-Abyad et al., 1993).
Effect of antagonistic organisms on chemical composition of tomato leaves: The results tabulated in (Tables 5 and 6) indicate that all antagonists organisms used in this study significantly affected chlorophyll a and b as well as carotenoids, phenols (free and conjugated) and sugars (reducing and non- reducing) compared with control. Data in Table 5 Illustrated that most of the tested treatment proved to be effective for increasing chlorophylls (a and b) and carotenoids content in leaves of tomato plants compared with control treatment. Concerning with phenols (free and conjugated) and sugars (reducing and non – reducing) in leaves of tomato plants as affected by tested treatments. Table 6 showed that all tested treatments caused significant increase in phenols and significant decrease in sugars content compared with control. The present results are in agreement with those of Khaled et al. (2000), they stated that leaves of resistant cultivar contained higher phenols and lower sugars. The present results are in conformity with those reported by Kuc (1964) who mentioned that phenolic compounds play a marked role in disease resistance and immunity of plant. Similar confirmation was reported by (khashaba, 1980), who stated that resistance of cotton to F. oxysporum f. sp. vasinifectum depends on the synthesis of phenolic compounds which are highly toxic to the pathogen. In conclusion, the present investigation showed the possibility of controlling wilt and early blight diseases of tomato by certain antagonistic microorganisms such as T. hamatum, T. harzianum, B. subtilis and P. fluorescens. These treatments also caused an increase in yield comparable to the untreated control. Three mechanisms in controlling soil-borne pathogens biological are proposed.
| a: | minimize the population of the pathogen in the soil by direct antagonism (Coolins and Papavizus, 1989). |
| b: | prevent the pathogen to infect the lost by competing in space or on nourishment (Ruppel et al.,1983). |
| c: | limiting disease development by altering the defense mechanisms in host tissues (Wells, 1988). |
REFERENCES
- El-shanshoury. A., H.E. Agwa, M.S. Osman and Y.H. El-Halmouch, 2000. Antifungal activities of Streptomyces anulatus, S. chromofuscue, S. albaduncus and Trichoderma viride against Corcospora baticola and Alternaria alternata causing sugar beet leaf spot. Bulletin of the Faculty of Science, Assiut University
- Ferreira, J.H.S., F.N. Matthee and A.C. Thomas, 1991. Biological control of Eutypa lota on grapevine by an antagonistic strain of Bacillus subtilis. Phytopathology, 81: 283-287.
Direct Link - Gurusiddaiah, S., D.M. Weller, A. Sarkar and R.J. Cook, 1986. Characterization of an antibiotic produced by a strain of Pseudomonas fluorescens inhibitory to Gaeumannomyces graminis var. tritici and Pythium spp. Antimicrob. Agents Chemother., 29: 488-495.
Direct Link - Papavizas, G.C., 1985. Trichoderma and Gliocladium: Biology, ecology and potential for biocontrol. Ann. Rev. Phytopathol., 23: 23-54.
Direct Link - Utkhede, R.S. and J.E. Rahe, 1980. Biological control of onion white rot. Soil Biol. Biochem., 12: 101-104.
CrossRef - Wolffhechel, H. and D.F. Jensen, 1992. Use of Trichoderma harzianum and Gliocladium vitens for the biological control of post-emergence damping-off and root rot of cucumbers caused by Pythium ultimum. J. Phytopathol., 136: 221-230.
CrossRefDirect Link