
A.U. R. Saljoqi
Department of Plant Protection, NWFP Agricultural University, Peshawar, Pakistan
H.F. van Emden
Department of Horticulture, School of Plant Sciences, The University of Reading,
Whiteknights, PO Box 221, Reading, RG6 6AS, UK
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 4 | Page No.: 386-393







ABSTRACT
In a field experiment the effects of Sumicidin (super) 5EC (fenitrothion), Metasystox EC25 (oxydemeton-methyl) and Tamaron SL600 (methamidophos), applied at different dosages, were evaluated against peach-potato aphid, Myzus persicae (Sulzer) and its parasitoid Aphidius matricariae Haliday on Cardinal and Desiree (respectively partially resistant and susceptible potato cultivars to M. persicae). Sumicidin (super) 5EC was found about 30% more effective in reducing aphid populations than the other insecticides tested. The highest doses of each insecticide caused maximum aphid mortality; in general aphid mortality appeared dose dependent. Almost all the higher and lower doses of the tested insecticides were about 19% more effective on Cardinal than on Desiree. The most significant result was the synergistic interaction at the lower doses with plant resistance, so that the same level of control was recorded with second highest dose on Cardinal as with the highest dose on Desiree. Also the same control level was achieved at the lowest dosage rate on Cardinal compared with the next higher dose on the Desiree. Sumicidin (super) 5EC was found least toxic to the parasitoid, A. matricariae in terms of percent parasitism, emergence of parasitoids and number of mature eggs in the emerging female parasitoids; increase of about 22, 67 and 47% respectively were found in parasitoid performance with Tamaron SL600 which was found comparatively highly toxic. The highest doses of all insecticides were found clearly toxic to the parasitoid. In general, effects on the parasitoid were dose dependent. Maximum yield was obtained from the second highest dose of Sumicidin (super) 5EC.
PDF Abstract XML References Citation
How to cite this article
A.U. R. Saljoqi and H.F. van Emden, 2003. Differential Susceptibilities of Peach-potato Aphid,
Myzus persicae (Sulzer) (Homoptera: Aphididae) and its Parasitoid
Aphidius matricariae Haliday (Hymenoptera; Aphidiidae) to Foliar
Insecticides on Partially Resistant and Susceptible Potato Cultivars. Pakistan Journal of Biological Sciences, 6: 386-393.
DOI: 10.3923/pjbs.2003.386.393
URL: https://scialert.net/abstract/?doi=pjbs.2003.386.393
DOI: 10.3923/pjbs.2003.386.393
URL: https://scialert.net/abstract/?doi=pjbs.2003.386.393
INTRODUCTION
Myzus persicae (Sulzer) is one of the most serious agricultural insect pests in the world (Van Emden et al., 1969; Mackauer and Way, 1976; Radcliffe, 1982). This species can feed on > 40 families of plants (Blackman and Eastop, 1984) and transmit over 100 viruses to plants belonging to different families such as Solanaceae, Cucurbitaceae, Leguminosae, Chenopodiaceae, Cruciferae, Compositae and Gramineae (Adams, 1946; Kennedy et al., 1962; Radcliffe and Lauer, 1966; Gibbs and Harrison, 1976; Behura, 1979). M. persicae is one of the most serious pests of potatoes in Pakistan (Shah, 1988). Infestation decrease yields by reducing plant growth (Mackauer and Way, 1976) and vectoring different virus diseases. Potato leafroll virus (PLRV) and potato virus Y (PVY) are the most important of these viruses limiting potato yield and quality (Banttari et al., 1993). Heavy infestation of this aphid can cause considerable damage to the potato crop by severely dwarfing and curling the leaflets and by dwarfing and spindling the tops. In extreme cases, the whole plant may be killed (Painter, 1951).
Disadvantages of heavy reliance on chemical pesticides to control M. persicae and other pests include problems of pest resistance, environmental and human health risks and the reduction in numbers of beneficial insects that might other wise help to control the populations (Metcalf, 1982). A major goal of IPM is to integrate multidisciplinary methodologies that are practical, effective, economical and protective of both public health and the environment (Smith, 1976). It does not attempt to eliminate pesticide usage, but rather to use the combination of different control methods to produce a stable and sustainable agroecosystem (Madder, 1986). A partially resistant variety can be used as an adjunct to other control measures and interact well with other components of IPM, i.e. chemical and biological control measures (Van Emden, 1987). Various studies have shown significant differences that insects reared on resistant plant cultivars are less tolerant to insecticides when compared with individuals reared on susceptible cultivars (Swingle, 1939; Chalfont and Brett, 1967; Selander et al., 1972; Wiseman et al., 1973; Kea et al., 1978; El-Refai et al., 1979; Heinrichs et al., 1984; Campbell and Wynne, 1985; Abro and Wright, 1989; Hinks and Spurr, 1989; Mohamed and van Emden, 1989; Ghidiu et al., 1990). The influence of host plants on susceptibility to insecticides may vary according to the insecticide used (Wood et al., 1981).
One of the major tools of IPM is to identify pesticides and practices that are more selective against the pests than against species useful for biological control (Georghiou, 1986). Unfortunately, most chemical pesticides used in agriculture are more injurious to natural enemies than the targeted pests (Sechser, 1981). In recent years there has been considerably interest in the development of programmes that assure a more compatible use of chemical and biological methods of pest control. Very few studies have dealt with their effects on parasitoids, especially aphid parasitoids (Horn, 1983; Mishra and Satpathy, 1985; Mullin and Croft, 1985; Van Emden, 1988). The toxic effects of a pesticide on the natural enemy especially on the parasitoids are very complex. They can cause acute mortality, but various sublethal effects may alter their reproductive potentially (Irving and Wyatt, 1973; Croft and Brown, 1975; O’Brien et al., 1985). In assessing the effects of insecticides on arthropod natural enemies, it is necessary to evaluate not only the direct mortality caused by contact or residual toxicity but more importantly any absence of such effects and also sub-lethal effects on emergence, survival, fecundity and predation/parasitism behavior if the insecticide impact is to be related to the total performance of natural enemies in the field (Croft, 1977).
Research work regarding the integrated management of M. persicae on the potato crop is very rare, especially in the field conditions. The information necessary to the compatibility of potato insect resistance with the appropriate active ingredient that favorably provided M. persicae control with the least adverse effects on the Aphidius mtricariae Haliday is lacking. There fore the present field study was conducted to ascertain a scheme for the integrated management of M. persicae, by investigating the effects of and the interaction between different pest management techniques i.e. insecticides, plant resistance and parasitoid. The objectives were to find out the most effective foliar insecticide, to determine minimum effective dose of each insecticide and to examine the interaction of partially resistant and susceptible potato cultivars with insecticides and their different dosage rates at different time intervals after application in relation to percent kill of M. persicae and effects on the percent parasitism, percent emergence and fecundity of the parasitoid and yield of the potato crop.
MATERIALS AND METHODS
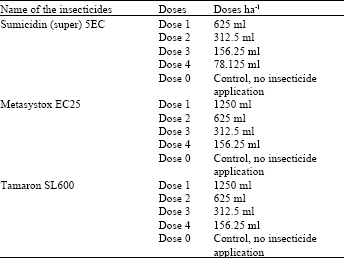
The experiment was conducted at the Agriculture Research Farm of NWFP Agricultural University Peshawar, Pakistan during spring season 1996. The partially resistant Cardinal and the susceptible Desiree cultivars were grown in a factorial randomized complete block design (RCBD) of 120 plots, representing thirty treatments (three active ingredients ( four doses + control) two potato cultivars) in four replications. Three foliar insecticides of two different groups, i.e. the pyrethroid Sumicidin (super) 5EC (fenitrothion) and the organophosphates Metasystox EC25 (oxydemeton-methyl) and Tamaron SL600 (methamidophos), were used. These insecticides were applied in four different doses (Table 1). Dose 1 was the recommended dose and doses 2, 3 and 4 were reduced dosage rates (½, 1/4 and 1/8 of the recommended dose, respectively) with dose 0 as the control, i.e. no insecticide application. Knapsack sprayer was used for the application of these insecticides. These insecticides were applied seven weeks after sowing the crop, when the aphid population was at its peak. The data recorded were: the number of aphids, percent parasitism of M. persicae, emergence rate and fecundity of the parasitoid and yield of potatoes. Natural parasitisation by A. matricariae was allowed to occur. The percent parasitism of M. persicae, emergence rate and fecundity of the parasitoids were recorded on the third day after spray application. The detail of data collection is given below.
M. persicae population estimate: The aphids were counted on the 1st, 2nd, 3rd, 10th and 17th day after spray application. These were counted on six tagged leaves, two each in the top, middle and lower region of four randomly selected plants, avoiding the border rows, from each plot. The data were recorded on the same leaves on the different sampling occasions (Singh and Verma, 1985; Trivedi and Misra, 1989).
Percent parasitism of M. persicae by A. matricariae: Percent parasitism was recorded by collecting 50 leaves (fixed number) from each plot. The mummies were removed and the remaining aphids were counted and kept in polythene bags (32x46 cm) still on the leaves with two filter papers (11.0 cm) for absorbing condensation. After five days, which would have allowed parasitoids to develop to 4 and 5th instar nymphs and adult aphids to form mummies, the number of mummies formed was counted and the percentage parasitism was calculated by the following formula:
 |
Emergence rate of the female parasitoid, A. matricariae from the mummified peach-potato aphid, M. persicae: Twenty mummies were collected from plants in each plot and placed in Petridishes, lined with filter papers. The Petridishes were covered and left in the laboratory. After ten days the number of emerged parasitoids, A. matricariae were recorded for each replicate and the percent adult emergence was calculated by using the following formula,
Fecundity of the parasitoid, A. matricariae: Fecundity of the female parasitoids was determined in terms of mature eggs present in the ovaries at emergence. Mummies were collected from each plot and placed in Petridishes. The newly emerged adult parasitoids were collected by means of a battery-operated aspirator from each Petridish and were provided with honey/water solution (30% on cotton balls). Sixteen adult female parasitoids were killed in 70% ethyl alcohol. The abdomen was removed and placed in glycerol on a microscope slide. The ovaries were extracted, using an insect mounting pin and placed in 1% acid fuchsin for 30 sec. The ovaries were then punctured under a low power stereo binocular microscope with an insect mounting pin to expel the eggs. These could then be counted under a high power microscope using a tally counter.
Yield: After hand harvesting the crop, the yield obtained was weighed separately for each plot in kg, which was then converted to tones per hectare.
Data analysis: The data were analyzed by analysis of variance of factorial randomized complete block Design (RCBD) with repeated measures ‘split-plot in time’ approach (Mead et al., 1993) using ‘MSTAT-C’ computer programme and the means were compared by using Least Significant Differences (LSD) (Steel and Torrie, 1960). For dose 0, a mean for the various data recorded was calculated separately for Cardinal and Desiree, combining all the untreated plots for each cultivar for each time interval. These plots were not included in the analysis of variance, to see only the effect of insecticides on M. persicae at different time intervals without having the control figures. The untreated means are given in the tables, so that the effect of the insecticide treatments can be seen.
RESULTS
Population of M. persicae: Table 2 shows the means for the interaction of cultivars ( insecticides ( post-treatment time intervals. On Desiree, Sumicidin (super) 5EC was found to be the most effective insecticide and remained superior throughout the experiment (P<0.05). Tamaron SL600 gave control, which overlapped with Metasystox EC25 statistically on all occasions (P>0.05), but recorded higher mean number of aphids from day 3 after application.
| Table 1: | Different dosage rates of the foliar insecticides |
 | |
| Table 2: | Effect of the interaction of potato varietiesxfoliar insecticidesxpost-treatment time intervals on the number of M. persicae, per six leaves of potato plants (means of 4 replicates) |
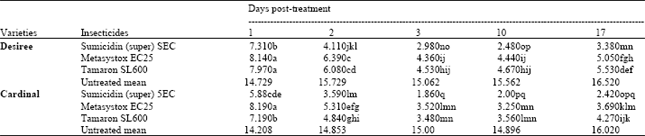 | |
| LSD for interaction = 0.590 (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control means) | |
The table also shows that maximum number of aphids was recorded on control throughout the experiment compared with all other treatments. A similar picture was seen on Cardinal. Sumicidin (super) 5EC again showed superiority on all sampling occasions over Metasystox EC25 and Tamaron SL600 (P<0.05). Tamaron SL600 gave better control than Metasystox EC25 on day 1, but later on as on Desiree, appeared less persistent. (Table 2).
Table 3 shows the interaction of cultivars ( doses. Highest number of aphids was recorded on control (on Desiree higher than Cardinal) compared with all other treatments. Cardinal was found superior and recorded a lower number of aphids at all dosage rates compared with Desiree (P<0.05), apart from the same performance of both cultivars at the highest dosage rate (P>0.05). The interaction lay in the progressive increase in number of aphids on Desiree as dose decreased (P<0.05), contrasted with no similar increase in number of aphids at dose 2 on Cardinal (P>0.05). The table also shows that the recommended dosage rate on the susceptible cultivar (Desiree) showed the same level of control as recorded from its half dosage rate on the partial resistant cultivar (Cardinal). Also the same control level was achieved at the lowest dosage rate on the partially resistant cultivar (Cardinal) compared with the next higher dose (dose 3) on the susceptible cultivar (Desiree).
Insecticides ( post treatment time intervals also interacted with dosage rates (Table 4). Again Sumicidin (super) 5EC gave the significantly best control throughout the experiment (P<0.05), apart from overlap with Tamaron SL600 on day 1 with the lowest doses (3, 4) (P>0.05). On day 1 Tamaron SL600 showed superiority at all doses over Metasystox EC25, apart from dose 2 where lower numbers of aphids were recorded for Metasystox EC25. On day 2 Tamaron SL600 was found significantly superior only at the lowest dose. On day 3 only at the highest dose, while on day 10 at the highest and also at the second highest doses now Metasystox EC25 showed superiority, which extended to the three lowest doses by day 17 (P<0.05). The table also shows that all treatments had significantly decreased the number of aphids compared with control at each time interval.
Aphids numbers were not transformed to log 10 (n+1) because variations between individual plants was not sufficient to require this.
Percent parasitism: Table 5 shows that maximum percent parasitism was recorded from the control compared with all other treatments. Sumicidin (super) 5EC was found consistently less toxic to parasitoid than Metasystox EC25 and Tamaron SL600 (P<0.05), which were statistically inseparable from each other (P>0.05).
| Table 3: | Effect of the interaction of potato varietiesxfoliar insecticides dosage rates on the number of M. persicae, per six leaves of potato plants (means of 4 replicates) |
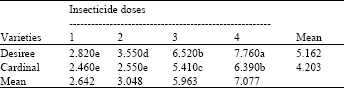 | |
Untreated mean for Desiree = 15.520, Untreated mean for Cardinal = 14.995, LSD for interaction = 0.375, LSD for doses = 0.266, (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control means) | |
| Table 4: | Effect of the interaction of the foliar insecticidesxdosesxpost-treatments time intervals on the number of M. persicae, per six leaves of potato plants (means of 4 replicates) |
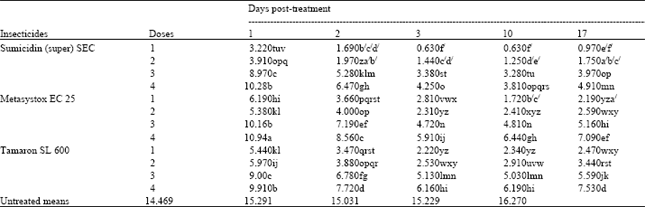 | |
| LSD for interaction = 0.446 (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control mean) | |
| Table 5: | Effect of foliar insecticides and their different dosage rates on the percent parasitism of M. persicae by its parasitoid (A. matricariae) in potatoes (means of 4 replicates) |
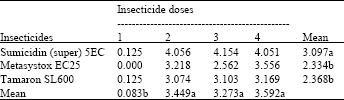 | |
Untreated mean = 8.110, LSD for insecticides = 0.535, LSD for doses = 0.617, (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control mean), Means followed by a different letter are significantly different from one another (P<0.05), using LSD test | |
| Table 6: | Effect of the interaction of foliar insecticides and their different dosage rates on the percent emergence of A. matricariae from mummified, M. persicae collected from Cardinal and Desiree potato varieties (means of 3 replicates) |
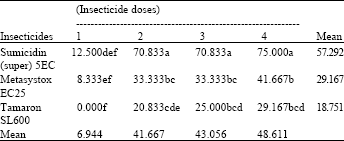 | |
Untreated mean = 90.276, LSD for interaction = 18.076, LSD for insecticides = 9.038, LSD for doses = 10.440, (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control mean) | |
| Table 7: | Effect of the interaction of foliar insecticides and their different dosage rates on the fecundity of the female parasitoid A. matricariae on potato plants (means of 4 replicates) |
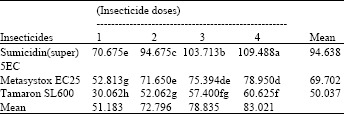 | |
Untreated mean = 150.945, LSD for interaction = 5.369, LSD for insecticides = 2.684, LSD for doses = 3.100, (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control mean) | |
| Table 8: | Effect of the interaction of foliar insecticides and their different dosage rates on the yield (t ha-1) of potato crop (means of 4 replicates) |
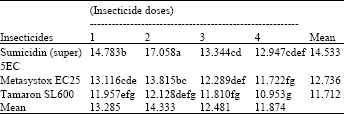 | |
Untreated mean = 10.390, LSD for interaction = 1.308, LSD for insecticides = 0.653, LSD for doses = 0.755, (An even smaller LSD would apply to any comparison with control means, in view of the larger number of replicates contributing to the control mean). Means followed by a different letter are significantly different from one another (P<0.05), using LSD test | |
Significantly reduced parasitism (P<0.05) was recorded by the application of dose 1 compared with the other doses, which were statistically similar to one another (P>0.05).
Emergence of A. matricariae: Table 6 presents the interaction of insecticides (dosage rates on the mean percent emergence of the A. matricariae. The table shows that parasitoids emerged well from the plants treated with the reduced doses of Sumicidin (super) 5EC, but that the highest dose was clearly damaging as it was for Metasystox EC25 and especially for Tamaron SL600. All the reduced doses of Metasystox EC25 and Tamaron SL600 also reduced parasitoid emergence. Highest percent emergence of the parasitoid was recorded from the control compared with all other treatments.
Fecundity of the parasitoid, A. matricariae: The means for the interaction of insecticides and their dosage rates are given in Table 7. The table shows that decreasing dosage rates of each insecticide increased the number of mature eggs recorded from the parasitoids. Maximum number of eggs was recorded from the female parasitoids emerging from the mummies collected from control. Sumicidin (super) 5EC was found superior at all its dosage rates and gave the maximum number of eggs in the female parasitoids, followed by Metasystox EC25 and Tamaron SL600 (P<0.05). The interaction lay in the progressive decline in parasitoid egg numbers as Sumicidin (super) 5EC dose increased, contrasted with the similar reduction in fecundity at the two highest doses for both Metasystox EC25 and Tamaron SL600.
Yield: The means for the interaction of insecticides x dosage rates are given in Table 8. The table shows that all treatments other than Tamaron SL600 dose 4 had increased yield compared with control. Sumicidin (super) 5EC was found superior and recorded the best yields at all dosage rates followed by Metasystox EC25 and Tamaron SL600. The lowest doses of Sumicidin (super) 5EC and Metasystox EC25 significantly depressed yield compared with doses 1 and 2. No differences on both doses were significant for Tamaron SL600, whereas for both Sumicidin (super) 5EC and Metasystox EC25 dose 2 significantly increased yield compared with doses 3 and 4. The peak yield was found at the second highest dose of Sumicidin (super) 5EC compared with the highest, perhaps due to losses of biocontrol and also suggesting some phytotoxicity.
DISCUSSION
The pyrethroid insecticide, Sumicidin (super) 5EC was found to be more effective in reducing aphid populations than the two organophosphate insecticides Metasystox EC25 and Tamaron SL600. These results agree with Singh and Verma (1985), who found that the pyrethroid insecticide (Sumicidin 20EC) was more effective against M. persicae than organophosphatic insecticide (Metasystox 25EC). The partially resistant cultivar (Cardinal) had fewer aphids than the susceptible cultivar (Desiree). Lowe (1975) also found lower number of M. persicae on resistant sugarbeet cultivar as compared with the susceptible cultivar. Mohamed and van Emden (1989) reported that M. persicae on a resistant Brussels sprout cultivar were not only smaller but also more sensitive to insecticides. The highest doses of each insecticide caused maximum aphid mortality in general, aphid mortality appeared dose dependent.
Fewer aphids were recorded after the 2nd day than before. The overall reduction of aphid population with all the tested insecticides was maximum on the 3rd day. Among the interaction of cultivars ( insecticides ( post-treatment time intervals, the minimum number of aphids was recorded on Cardinal on the 3rd and 10th day after the Sumicidin (super) 5EC application. It was also observed that the toxicity of the insecticides decreased with the passage of time. The aphid numbers did not appear to increase much in untreated plots during the experiment. This may be because of the presence of biological control agents (both larvae and adults of coccinellids and syrphids were observed in the untreated plots).
Percent parasitism, emergence of parasitoids, the number of mature eggs extracted from female parasitoids were depressed by all insecticides, but less so by the application of the pyrethroid Sumicidin (super) 5EC, which was significantly better in all these aspects than the organophosphate insecticides (Metasystox EC25 and Tamaron SL600). Regarding dosage rates, a better percent parasitism, emergence of the parasitoid occurred at reduced doses for all insecticides, which were statistically similar to one another but all significantly better than their highest doses. The highest emergence was recorded with all the reduced doses of Sumicidin (super) 5EC, which were statistically similar (P>0.05). The lowest emergence was recorded with the application of the highest dose of Tamaron SL600. The maximum number of mature eggs was found with the lowest dose of Sumicidin (super) 5EC. These results corroborate those of Abo El-Ghar and El-Sayed (1992) who tested five different insecticides against the cabbage aphid Brevicoryne. brassicae (L.) and its associated parasitoid Diaeretiella. rapae (M’Intosh). They reported that the pyrethroids were preferable to the organophosphate insecticides, because they had minimal adverse effects on the emergence of parasitoids from mummified aphids and their subsequent survival, longevity and fecundity, when applied at rates a little below the recommended field rates. Hsieh and Allen (1986), with both organophosphate and pyrethroid insecticides, found that increased doses of each insecticide reduced fecundity. They also reported that the organophosphate insecticides were more active and suppressed the fecundity of D. rapae at most doses when compared with pyrethroids. El-Sayed and Abo El-Ghar (1989) reported that increased doses of organophosphate, pyrethroid and carbamate insecticides reduced fecundity of D. rapae. They also reported that parasitoid fecundity was better with pyrethroid than with organophosphate insecticides.
The partially aphid-resistant Cardinal showed a higher yield both in untreated and insecticide treated plots than Desiree. In the interaction of insecticides and their dosage rates, maximum yield was recorded with dose 2 of Sumicidin (super) 5EC.
It was concluded that the recommended dose of each insecticide caused maximum aphid mortality, but on the other hand caused significant reduction in the percent parasitism, emergence and fecundity of the parasitoids. Dose 2 of all insecticides seemed to provide a satisfactory level of control on the Cardinal, because it reduced the number of aphids sufficiently and also showed good percent parasitism, emergence, fecundity of the parasitoids and yield. Sumicidin (super) 5EC dose 2 gave the best results overall. Although we recorded good results in the percent parasitism, emergence and fecundity of the parasitoids for dose, 3 and dose 4 of all the insecticides, we noted an increased number of aphids due to the very reduced dosage rate and also recorded a low yield as compared with the dose 2.
ACKNOWLEDGMENTS
We thank the statistics advisory of The University of Reading for helping us in the data analysis. We are also grateful to the World Bank assisted Agricultural Research Project-II for financial support.
REFERENCES
- Abo El-Ghar, G.E.S. and A.M. El-Sayed, 1992. Long-term effects of insecticides on Diaeretiella rapae (M'Intosh), a parasite of the cabbage aphid. Pesti. Sci., 36: 109-114.
CrossRefDirect Link - El-Sayed, A.M. and G.E.S. Abo El-Ghar, 1989. Selective toxicity of organophosphorus insecticides to the cabbage aphid, Brevicoryne brassicae (Homoptera: Aphididae), and its parasitoid, Diaeretiella rapae (Hymenoptera: Aphidiidae). Zeitschrift fur Pflanzenkrankheiten und Pflanzenschutz, 96: 281-288.
Direct Link