
Habib-ur-Rehman Athar
Institute of Pure and Applied Biology, Bahauddin Zakariya University, Mullan, Pakistan
Seema Mahmood
Institute of Pure and Applied Biology, Bahauddin Zakariya University, Mullan, Pakistan
Rubina Shaheen
Institute of Pure and Applied Biology, Bahauddin Zakariya University, Mullan, Pakistan
Muhammad Ashraf
Department of Botany, University of Agriculture, Faisalabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 4 | Page No.: 317-321







ABSTRACT
Two populations of Cynodon dactylon (Linn.) Pers., from contrasting habitats, were assessed for waterlogging tolerance. Tillers from each population were subjected to 3 (control), 18, 36, 54, 72 and 90 mg l-1 Fe as Fe-EDTA in Rorison's nutrient solution for 21 days under controlled conditions. The results showed that addition of Fe to the growth medium had caused a significant inhibitory effect on fresh and dry matter of both grass populations. However, population from non-waterlogged habitat produced slightly greater shoot biomass than the waterlogged population in supplemented nutrient medium containing iron. Shoot K+ contents were significantly lower in waterlogged population at all external Fe regimes whilst, the two populations were not different for Ca2+ or Mg2+ contents of shoots. The waterlogged population accumulated significantly greater concentration of Mn in shoots. Likewise, individual plants sampled from waterlogged site exhibited greater N contents in their shoots as compared to the normal population. However, shoot P contents showed a non- significant decline in both populations under varying concentrations of iron. The normal population appeared to be more susceptible with respect to amino acids accumulation at higher concentrations of Fe. On the other hand, protein contents in shoots increased significantly in the normal populations at varying Fe regimes. It was concluded that the lack of tolerance to the excessive amounts of Fe in the waterlogged population was probably due to the absence adaptive mechanism, lack of suitable selection pressure and insufficient time for sorting of genotypes into that habitat.
PDF Abstract XML References Citation
How to cite this article
Habib-ur-Rehman Athar, Seema Mahmood, Rubina Shaheen and Muhammad Ashraf, 2003. Water Logging Tolerance in Populations of Cynodon dactylon from Contrasting Habitats. Pakistan Journal of Biological Sciences, 6: 317-321.
DOI: 10.3923/pjbs.2003.317.321
URL: https://scialert.net/abstract/?doi=pjbs.2003.317.321
DOI: 10.3923/pjbs.2003.317.321
URL: https://scialert.net/abstract/?doi=pjbs.2003.317.321
INTRODUCTION
In waterlogged habitats, surface layers of soil are saturated with water. Therefore, the spaces between the soil particles are occupied by water instead of air. Poor soil aeration associated with excessive moisture influence plant growth in various ways. Many physiological and morphological changes; reduced photosynthesis, epinasty, leaf abscission, chlorosis, decreased stem and root growth poor yield are manifestations of excessive moisture stress (Batzil and Dawson, 1997). But the most drastic change includes the altered pattern of amino acids and protein accumulation under stress conditions (Chang et al., 2000). The changes in soil characteristics involve low rate of oxygen diffusion, lowering of redox potential besides iron and manganese reduction. Moreover, carbon dioxide, ethylene, manganese and iron may accumulate in concentrations that are potentially toxic to plants (Ponnamperuma, 1984).
The most conspicuous chemical change under such conditions is the reduction of Fe3+ to Fe2+, which is the soluble form of iron and is an outcome of anaerobic bacterial activity (Yoshida, 1978). Both Fe and Mn in their reduced forms are very soluble and consequently, their enhanced uptake may cause toxicity symptoms in plants. Similarly, excessive iron can hinder the uptake of other nutrients, which results in the development of deficiency symptoms. Those plant species that inhabit waterlogged soil may use some protective mechanisms to cope with high concentrations of soluble forms of Fe and Mn (Armstrong, 1982; Talbot et al., 1987). Several workers have drawn parallels between waterlogging tolerance, uptake of iron and manganese under flooded conditions. Moreover, mineral nutrient status, amino acids and protein accumulation are other important aspects of plant species with differential waterlogging tolerance (Jones, 1971; Talbot et al., 1987).
Therefore, the present study was conducted to establish a relationship between uptake of reduced forms of iron and waterlogging tolerance or sensitivity in populations of Cynodon dactylon (Linn.) Pers. In addition, mineral nutrient status, accumulation of amino acids and proteins were also assessed in these grass populations in response to varying concentrations of iron supplied to the growth medium.
MATERIALS AND METHODS
A good collection (200-300 tillers) was made from two populations of C. dactylon during May 1998. The first population was referred to as normal since it was collected from a site with normal irrigation, whereas the other was sampled from a site with intermittent flooding and regarded as waterlogged population. These two sites were located at the Botanic Garden, Bahauddin Zakariya University, Multan.
All tillers from both populations were transplanted individually into plastic pots (internal diameter 18 cm) containing normal potting compost. Plants were grown for 10 weeks under glasshouse conditions and irrigated with normal tap water. Then 10 tillers of uniform size were separated from these pots and shifted to 500 ml plastic beakers containing nutrient solution (Hewitt, 1966) under non-aerated conditions. The tillers were allowed to establish for 5 days before the imposition of iron treatments. Fe-EDTA salt was used to prepare Fe solutions of varying concentrations. The six iron treatments were control (3 mg l-1 iron in nutrient solution), 18, 36, 56, 72 and 90 mg l-1.
The experiment was conducted for 21 days in the growth room at 26±3°C, 52% Rh. and 36 W m-2 light intensity. Treatment solutions in the beakers were changed after every seven days in order to maintain nutrient levels. The solutions were not aerated so that simulated waterlogged condition can be developed. Then after 21 days plant biomass production was determined and shoots were analyzed for the following physiological parameters.
Mineral nutrients: N, P, K, Ca, Mg, Fe Mn in shoots were determined by the methods described by Allen et al. (1986). K contents were determined with a flame photometer (Jenway PFP7), Ca, Mg, Fe, Mn and P with an atomic absorption spectrophotometer (Perkin Elmer Analyst 100) while, total N was estimated by titration method.
Soluble proteins and free amino acids: Soluble proteins and free amino acids were estimated as described by Lowry et al. (1951). Shoot proteins were extracted in sodium phosphate buffer (pH = 7) followed by the addition of folinphenol reagent. The optical densities of the reaction mixtures were read at 620 nm using spectrophotometer (Hitachi, U2000). For amino acids determination, ninhydrin solution was added to the buffer extracts and then optical densities of the coloured solutions were read at 570 nm. Total soluble proteins or free amino acids were estimated as follows:
| Where, | ||
| O.D | = | Optical density of sample |
| V | = | Volume of the sample |
| D.F | = | Dilution factor Statistical analysis |
The mean values for all parameters were calculated. The data for fresh and dry matter and for physiological parameters were subjected to a two-way analysis of variance using COSTAT (Cohort Software, Berkeley, California). Thus, main effects of iron treatments and differences between populations were assessed. Least significance difference (LSD) between means was determined following Snedecor and Cochran (1980).
RESULTS
The mean values for fresh weight of shoots of two populations of C. dactylon from contrasting habitats (Fig. 1) showed that increasing concentrations of iron in the rooting medium had an inhibitory effect on shoot fresh weights (P< 0.001). Populations also differed significantly for this response. Since, the populations differed significantly at the control treatment, it was not legitimate to compare the data for mean shoot weight, therefore, the data were transformed to percent of control. The two higher concentrations of Fe (72 and 90 mg l-1) proved to be considerably inhibitory for shoot fresh weight in both populations but a more profound effect was observed in the population collected from waterlogged site (Fig. 1).
Mean data for shoot dry weights also indicated significant (P< 0.01) effects of external Fe regimes and marked difference between populations (P< 0.001). The normal population was slightly better for shoot fresh matter as well as for shoot dry weight than the waterlogged population (Fig. 1).
It is evident from mean values that nitrogen contents increased consistently in the waterlogged population when Fe levels were raised in the growth medium but, by contrast, in the normal population shoot nitrogen remained unchanged at all external Fe applications (Fig. 2). There was a significant effect of external Fe regimes on shoot nitrogen (P< 0.05) but overall differences between populations did not become evident for this response. Shoot phosphorous decreased consistently in both populations with the increase in external Fe (Fig. 2).
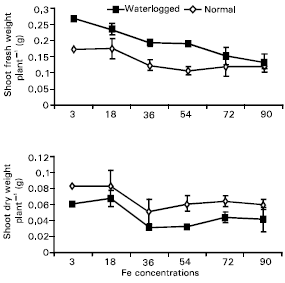 | |
| Fig. 1: | Mean values (± S.E) for fresh and dry weight (g) of shoots of two populations of Cynodon dactylon from contrasting habits grown for 21 days under different concentrations of Fe in Rorison’s nutrient solution |
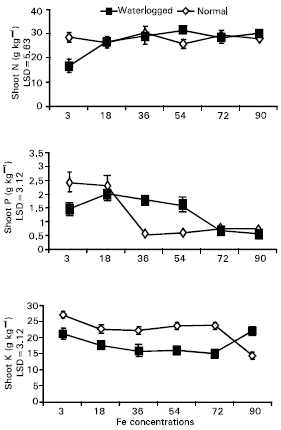 | |
| Fig. 2: | Mean values (± S.E) for N, P, and K contents (g kg -1 dry weight in shoots of two populations of Cynodon dactylon from contrasting habits grown for 21 days under different concentrations of Fe in Rorison’s nutrient solution (LSD at 5 %) |
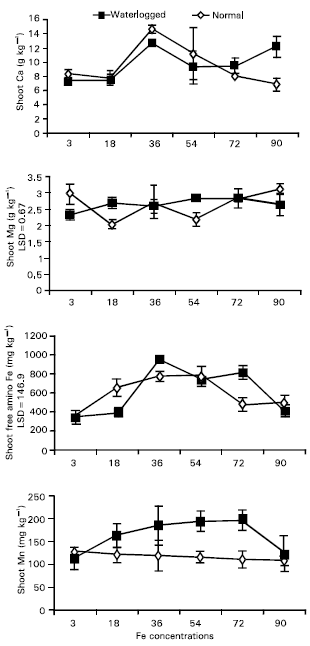 | |
| Fig. 3: | Mean values (± S.E) for Ca, Mg (g kg-1 dry weight), Fe and Mn (mg kg-1 dry weight) contents in shoots of two populations of Cynodon dactylon from contrasting habits grown for 21 days under different concentrations of Fe in Rorison’s nutrient solution (LSD at 5%) |
Likewise, K+ concentration in shoots (Fig. 2) decreased significantly (P< 0.001) in both populations but the pattern of decrease in K was not consistent in the waterlogged population. In general, waterlogged population had lower K in shoots than the non-waterlogged population at all external Fe regimes except at the highest Fe level (90 mg l-1) where the reverse was true. No consistent pattern was observed for increase or decrease in shoot Ca2+ (Fig. 3).
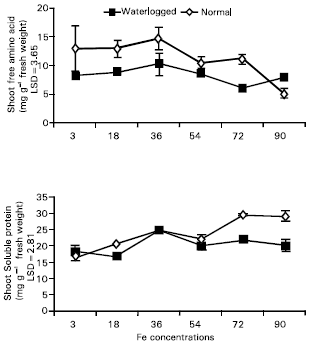 | |
| Fig. 4: | Mean values (± S.E) for three amino acid and soluble proteins (mg g-1 fresh weight) in shoots of two populations of Cynodon dactylon from contrasting habits grown for 21 days under different concentrations of Fe in Rorison’s nutrient solution (LSD at 5%) |
On the other hand, there was no significant effect of varying concentrations of Fe on Mg2+ concentration in shoots.
Fe concentration in shoots increased significantly (P< 0.001) with an increase in the external Fe regimes (up to 54 mg l-1) in non-waterlogged population but hereafter shoot February 27, 2003Fe concentration was as much as that at control in both populations (Fig. 3). Therefore, differences between populations for Fe content were not consistent. Waterlogged population has accumulated greater concentrations of Mn in shoots than the normal population (Fig. 3).
Fe regimes had a significant (P< 0.001) effect on free amino acids in shoots of both populations of C. dactylon. A drastic decrease in the shoot free amino acids was found in the normal population at elevated levels of external Fe. Whereas, the amount of free amino acids remained unaffected in the waterlogged population (Fig. 4).
The presence of Fe in the rooting medium had a significant influence on shoot soluble proteins (P< 0.001) and populations also differed significantly for this biochemical trait. A more profound change in shoot soluble proteins was observed in the normal population with increasing concentrations of Fe. Although a rise in protein contents was also observed at iron concentration of 36 mg l-1 in the waterlogged populations but this increase was found to be statistically non significant (Fig. 4).
DISCUSSION
Fresh and dry biomass production is an important trait of grass species. Shoot biomass data clearly depicted that population of C. dactylon collected from the waterlogged site had not shown tolerance to excessive concentrations of Fe and produced lower biomass. The evolution of stress tolerance mainly depends on the presence of appropriate gene pool, sufficient time, as well as on the existence of suitable selection pressures (Bradshaw, 1984). The populations of C. dactylon from contrasting habitats were found to be undifferentiated for waterlogging tolerance, when they were exposed to elevated levels of Fe. Though, rapid population differentiation in response to stress environment but the pace and efficiency of micro-evolution vary considerably in grass species. For example, in the presence of strong selection pressures, Agrostis stolonifera developed tolerance within four years of exposure to copper (Wu et al., 1975) and Anthoxanthum odoratum within six years of lime application (Snaydon and Davis, 1972). Hence, for this study it can be assumed that the force of natural selection had not yet worked within this short span of time.
While summing up the results for shoot N, P, K, Ca Mg it is evident that shoot K decreased significantly in both populations due to excessive amount of iron in the rooting medium. Therefore, these results supports the findings of Hocking et al. (1987) who indicated decline in K contents of shoot in cotton under flooded conditions. Similarly the uptake of other nutrients was also influenced under oxygen deficient conditions. Morard and Silvestre (1996) also reported that oxygen deprivation disturbed the uptake of mineral nutrients. Thus, the results reported here are in conformity with the study of these workers.
The contrasting populations did not show any dissimilarity with respect to their shoot Fe and Mn contents. However, shoot Mn contents were slightly higher in population sampled from waterlogged site than its respective normal population. Therefore, greater uptake of Mn cannot be attributed to flooding tolerance. Talbot et al. (1987) have suggested that oxidizing capacity of roots hinders the uptake of Fe and Mn ions thus, facilitates water logging tolerance but this study has not revealed such mechanism.
It can be assumed that flooding has not caused any additional accumulation of stress amino acids or proteins regardless of continuous N uptake. Hence, these results are in agreement with Bacanamwo and Purcell (1999) who reported non-adaptation of soybean to flooding because of unchanged pattern of amino acids and protein synthesis in relation to N accumulation under waterlogged conditions.
This study showed that the population of C. dactylon that was sampled from a waterlogged site did not show optima of flooding tolerance. It is concluded that non- adaptation to toxic levels of Fe under waterlogged conditions has not evolved in the species. It is presumably due to lack adaptive mechanism, absence of strong selection pressure and insufficient time for sorting of genotypes into that particular environment.
REFERENCES
- Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Morard, P. and J. Silvestre, 1996. Plant injury due to oxygen deficiency in the root environment of soilless culture: A review. Plant Soil, 184: 243-254.
CrossRefDirect Link