
Sofia Alvi
PCSIR Laboratorie Complex Off. University Road, Karachi-75280, Pakistan,
Rashida Perveen
Department of Chemistry, University of Karachi, Karachi�25270, Pakistan
Iftikhar Imam Naqvi
Department of Chemistry, University of Karachi, Karachi�25270, Pakistan
S. Shahid Shaukat
Department of Botany, University of Karachi, Karachi�25270, Pakistan
Pakistan Journal of Biological Sciences
Year: 2003 | Volume: 6 | Issue: 3 | Page No.: 249-251







ABSTRACT
Effect of atrazine was determined on the absorption and accumulation of phosphorous (P32 ) in bean plants and results have been explained in relation to other physiological processes. Atrazine seemed to be highly toxic to the seedlings at all concentrations used. The injury caused by the herbicide increased with its concentration. It was found to decrease the contents of chlorophyll a, chlorophyll b and carbohydrates, which reflects its effectiveness as a photosynthetic inhibitor. Potassium and protein contents were decreased significantly at 10 to 100 ppm atrazine. The uptake of phosphorus was also recorded and it declined with time. The accumulation of P32 was inhibited at 10 to100 ppm concentrations of atrazine. Some growth stimulating effects were also observed at 5 ppm level.
PDF Abstract XML References Citation
How to cite this article
Sofia Alvi, Rashida Perveen, Iftikhar Imam Naqvi and S. Shahid Shaukat, 2003. Effect of Atrazine on Absorption and Translocation of P32, Chlorophyll, Carbohydrate,
Protein and Potassium Contents in Bean Vigna radiata (L.) Wilczek. Pakistan Journal of Biological Sciences, 6: 249-251.
DOI: 10.3923/pjbs.2003.249.251
URL: https://scialert.net/abstract/?doi=pjbs.2003.249.251
DOI: 10.3923/pjbs.2003.249.251
URL: https://scialert.net/abstract/?doi=pjbs.2003.249.251
INTRODUCTION
Atrazine is a prominent triazine and is one of the most widely used herbicides in the world of agriculture. It is a selective pre-emergence herbicide for the control of many grasses, broad leaf weeds in maize, sorghum and sugarcane crops, rangeland and forest. It increases the yield of crop (Shah et al., 2000) and also used as a nonselective herbicide in non crop areas and in minimal or no tillage programmes in maize and sorghum. Many vegetable crops are sensitive to atrazine. Chemically it is 2-choloro-4- (ethyl amino) -6- isopropyl-s-triazine and is available as wettable powder.
Atrazine is purely soil applied herbicide. However, it also has foliar activity when applied with an additive. It is apoplastically translocated in plant. Extensive research conducted on the absorption and translocation indicated that atrazine is taken up by the roots and moves rapidly to the top of the apoplast. It concentrates first in veins, then in interveinal areas and finally in the margins of the leaf. There is no apparent symplastic movement out of the leaf (Ross and Lembi, 1985).
Atrazine toxicity is primarily due to the effects on photosynthetic processes such as Hill reaction and its associated non-cyclic photophosphorylation. Atrazine concentration greater than 0.05 μm inhibited the protoplast O2 evolution (McCloskey et al., 1990). It is also found that atrazine accelerated the vacuolation in cell and disintegration of chloroplast in leaves of the treated plants. Giannopolitis and Ayer (1978) reported atrazine-based inhibition of photosynthetic electron transport and acceleration of photo-oxidation. Boydstone et al. (1986) discovered that atrazine and tridiphane combination impaired the photosynthetic rates more effectively. The triazine herbicides affect the plant growth by inhibition of photosynthesis, it might change the performance of the crop and its nutrition (Hence, 1985). The present investigation aimed to study 1) the effect of atrazine concentration on the absorption and accumulation of phosphorous in bean plant, 2), to determine the influence on potassium uptake, carbohydrate, protein and chlorophyll contents.
MATERIALS AND METHODS
The pot experiment was conducted in a growth chamber (30°C day and 25°C night) for 15 days in September 2001. The seeds of mungbean were washed and soaked in distilled water for 4 h and washed with 0.3% calcium hypochlorite. The broken and damaged seeds were discarded. Seeds were germinated in half strength Hoagland solution containing 0, 5, 10, 50 and 100 ppm atrazine (80% wettable powder), Few drops of tween 20 were used as dispersing agent (Chapman, 1976). Plants were analyzed after 15 days of germination. Chlorophyll a and b contents in shoot portion were determined in extract of shoot in 98% acetone spectrophotometrically by recording the absorbance at 663 and 645 nm, respectively (Ahmed et al., 1985). Carbohydrate contents were analyzed in protein free filtrate extract of plant in water using anthrone reagent by spectrophotometer at 620 nm (Tandon, 1993). Proteins were estimated by Folin Cicalteau phenol reagent in water extract of bean plant and analyzed spectrophotometrically at 650 nm (Lowry et al., 1951). Potassium was estimated in dry ashed samples (Chapman, 1976) by flame photometry. In order to determine the effect of phosphorous accumulation in the atrazine treated plants, 8 plants were selected at random and were placed in tubes containing activity 10 μci P32. The tubes were left for about 1.5 h, subsequently the plants were washed properly and were transferred to their respective herbicide solution. To obtain time scan data, plants were analyzed after 4, 8, 12 and 24 h. Plants were cut into root, shoot and leaves. One part was crushed in a small quantity of ethanol to get a homogeneous distribution of activity, dried and number of counts were determined on a G.M. counter (The Nucleus, USA).
RESULTS AND DISCUSSION
Atrazine seemed to be highly toxic to the seedlings of bean plants having concentrations 10 to 100 ppm. The height of the plant was increased at 5 ppm atrazine but it decreased significantly up to 100 ppm. Shoot length was more affected as compared with root (Fig. 1a). High concentration of atrazine is known to reduce germination percentage and shoot and root growth (Shaukat and Soni 1974; Shaukat, 1976; Burhan and Shaukat, 2000). The contents of both chlorophyll a and b were decreased in plants treated with 10 to 100 ppm concentration of atrazine (Fig. 1b), but chlorophyll a and b did not change significantly at 5 ppm level (Fig. 1b). Atrazine has been reported as a potent photosynthetic inhibitor. This may in part be due to reduction of chlorophyll synthesis in atrazine treated plants. Carbohydrate contents were decreased from 10 to 100 ppm atrazine level. At 5 ppm level, the contents were about the same as for control (Fig. 1c). The carbohydrates are synthesized in the process of photosynthesis, their levels are also affected due to reduction in chlorophyll contents. Potassium uptake was significantly decreased in plants treated with 10 ppm or more of atrazine, but it was increased at 5 ppm concentration of atrazine (Fig. 1d). Uptake of ions like potassium and phosphate presumably decreased due to the damage of vascular tissues and lowered energy levels in atrazine treated plants (Shaukat et al., 1975; Hence, 1985). The uptake of potassium and phosphate ions in triazine (including atrazine) treated plants has also been reported by Ebert and Dumford (1976). Potassium is associated in cell membrane, ionic balance, opening of stomatas and other plant movements as well enzymes and protein synthesis (Hale and Orcutt, 1987). Low levels of proteins at 10 to 100 ppm concentration of atrazine may be associated with this fact (Fig. 1e). Proteins in plants treated with 5 ppm were increased (Fig. 1e), which may be due the stimulatory effect at this concentration of atrazine.
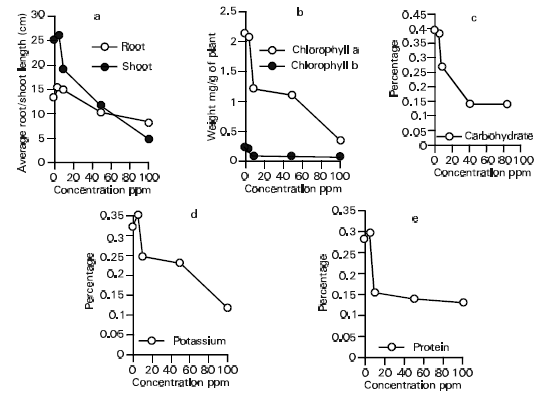 | |
| Fig. 1: | Effect of atrazine on (a) root and shoot length (b) chlorophyll a and b © carbohydrates (d) potassium and (e) protein contents |
| Table 1: | Effect of atrazine on accumulation of phosphorous at different time period at different concentrations |
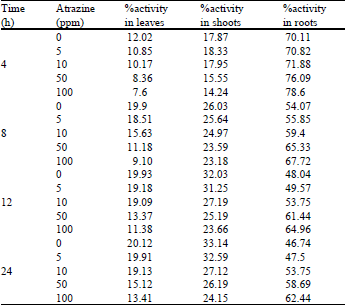 | |
The upward movement of P32 in atrazine treated plants was very slow. At higher levels of atrazine (50 to 100 ppm), after 24 h only 13.4% activity was transferred to leaves as compared to 20% for control (Table 1). At low levels of atrazine (5 to 10 ppm ), the rate of accumulation was initially slow (1% and 15% as compared to 12 and 19% for control after 4 and 8 h, respectively), but after 24 h the transfer rate was the same as for controls and 19.9%, respectively (Table 1). This was presumably because of lower energy levels available in the beginning for the active transport. The accumulation of P32 in shoot was low from 10 to 100 ppm (Table 1) and most of the P32 remained in roots after 24 h (Table 1). The 5 ppm level showed same time scan pattern as for control for the shoots and roots (Table 1).
In general injury caused by herbicide application to the root increased with the increase in herbicide concentration in the external solution, which in turn was probably associated with the increase in herbicide uptake by the roots (Shaukat and Soni, 1974).
Atrazine reduced root and shoot length of bean plant presumably is the result of inhibition of photosynthesis. Carbohydrate and protein contents declined as a result of reduced synthesis, while uptake of potassium and P32 was impeded because of reduced energy levels and possibly due to damage of root or vascular tissues. Increase in root and shoot length at 5 ppm concentration of atrazine may be associated with increase in potassium and protein contents.
ACKNOWLEDGMENT
We are thankful to pinstect for providing us P32 free of cost. We are also acknowledged to Dean Science for financial support.
REFERENCES
- Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Shaukat, S.S., 1976. Effect of simazine, atrazine and 2,4-D on respiration rate, sugar level and amylase activity during germination of Pinus nigra var. calabrica Schneid. Pak. J. Bot., 8: 37-45.
Direct Link